Decades of experiments in the area of diet, showed the immense benefits of a fully unprocessed raw food diet and regulated by our senses of smell and taste. This instinctive diet, once we learn to trust our instincts repressed by a lifetime of culinary habits, allows our body to reach opitmal health with minimal efforts.
Time showed consequences to be so numerous and far-reaching, as to liken the advent of cooking deep in prehistory to the Fall of the garden of Eden or similar myths throughout the world.
The first and most obvious observations, was the responsability of cooking for most bacterial, viral and degenerative diseases. In fact our very understanding of disease is wrong, as represented by the fallacious Germ theory whose own creator understood that the terrain of the body is what plays a crucial role in the progress of disease. However, with a certain amount of food discipline and a fairly broad range of healthy foods, a very strict discipline and obedience to the sense of smell (letting the body determine its needs), we can use our body as it was meant, once again.
It is possible to awaken our eating instinct, which all animals have, and bring about the near complete elimination of disease, especially as a life-threatening phenomena. Strength, agility, recovery time, mental acuity, all stats improve samewise on their own, fairly quickly, in proportion to the quality of your diet (and secondarily of your genetics). Access to the food range as close as can be to that of the great apes (whose needs are near-identical to our own) also proved absolutely essential.
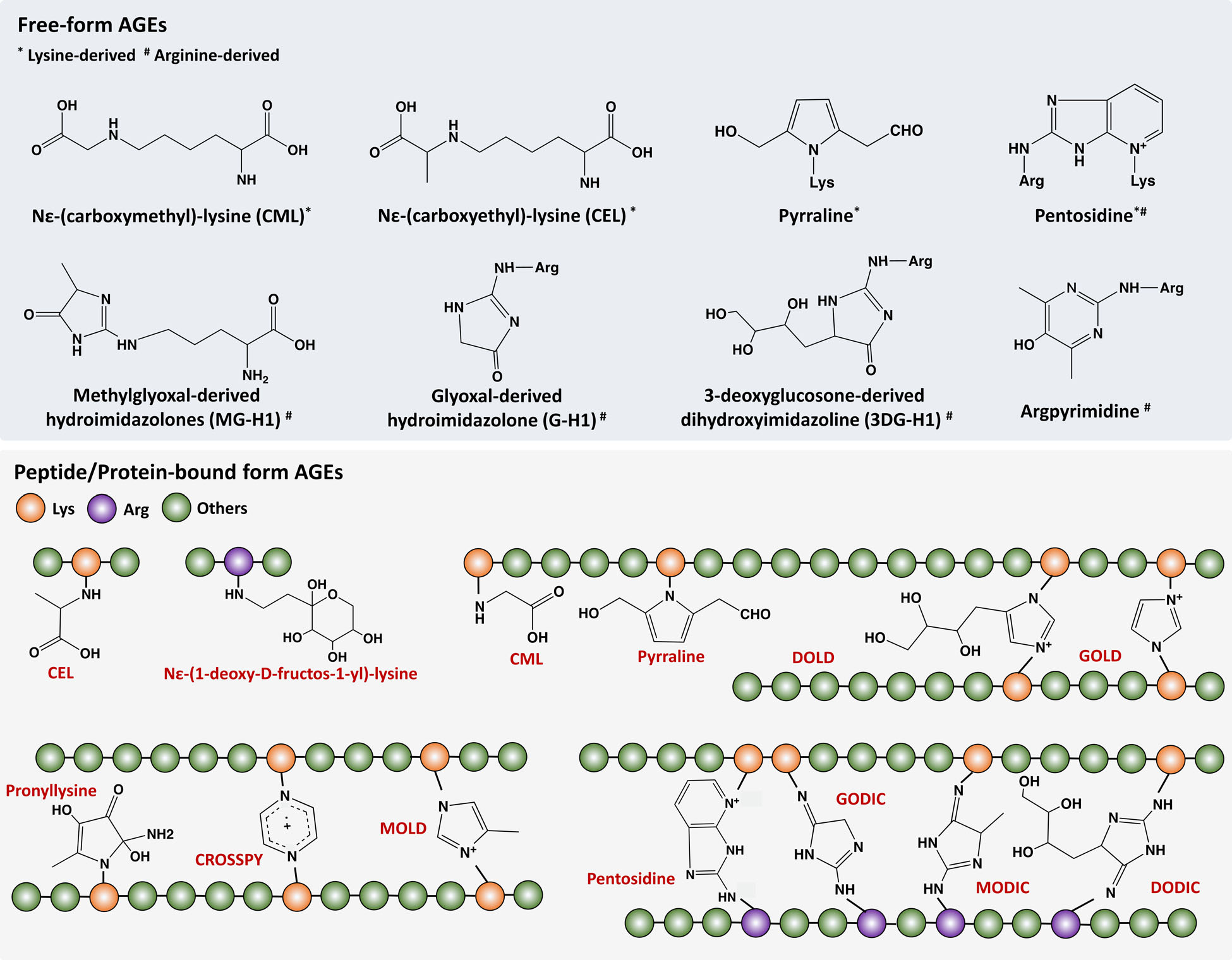
The main reason is the AGEs (and ALE for lipids) are classes of very specific molecules, thousands of times more abundant in cooked food and associated with disorders as varied as cancer, diabetes, organ failure, cognitive decline with and without Alzeimer disease, with genetics markedly playing no role. AGEs are the results of a chaotic blending of sugar chains and proteins and we know of their key role in inflammations (making for a much higher pain sensibility in humans than in wild animals), and last but not least the aging process. Except for methylglyoxal10,12,13—a product of normal metabolism—all other AGEs do not exist naturally in our body and must derive from food… that is, processed food.
Evidence suggests that this impact of dietary AGEs on different signalling pathways contribute to the onset of organ damage throughout on the body, especially muscles and liver and brain, affecting both metabolic control and global health.
Detecting, quantifying and studying AGEs in live subjects, has only been possible very recently, though precision sufficient for really conclusive studies still require mass spectrometry for the simple reason that it is the only way to study molecules whose structure we just don’t precisely. And considering their untold diversity, low concentration and possible intracellular location, the need for a bulky (and expensive) device such as a mass spectrometer definitely hinders the understanding of AGEs, their role and ubiquity… supposing scientists had the drive to invest in this research in the first place. So we see that the study of AGE, even the means for it, have not grown out of its diapers.
These molecules are produced during cooking at a rate exceeding several thousands times if not more than what is found in nature (outside forest fires of course, though they usually burn trees to a crisp instead of cooking them). Yet despite several findings indicating their major role in health, nothing even remotely of a corresponding scale was undertaken to curtail the issue. Even the bloodiest dictator could never fight against his people’s habit of cooking… We will see what consequences this constant chemical onslaught has been tampering with the brain and disrupting its higher functions for tens of millenia (increasingly so), twisting the development of culture since then. we will also see the omnipresent effects on health in general, creating what is in effect is a false, feeble human condition, a fallen nature.
But from a certain threshold of pollution, a bit more or less of it can decently be expected to predict much. Once cooked, to tell who will die when or how becomes like predicting which tree will fall first in the next hurricane. Sanguine concentration (of AGEs) is mostly of a qualitative indicator and can not be naively used to rank people for the likelihood of diseases, let alone patients for the progression of specific diseases in very limited conditions.
For one, those are chaotic, pathological processes, hence to expect a nice and simple linear correlation flies in the face of common sense. Besides, one should not forget the presence of multifarious compensatory mechanisms, the impact of genetics and other environmental factors, all things hard to intuit, and impossible to control for.
Secondly, because the body innately recognize a subset of all AGEs as normal physiological junkthere is no reason for all AGE to cause the same trouble, when we consider that the body is used to deal with some (the simpler kinds, one could assume) as
Thirdly, many use the absence of a convenient linear relationship as the proof of innocuity of a traditional, AGE-rich diet, as in plainly stated in the abstract of their studies.
As should be expected, the levels of thermal process contaminants in foods does not reflect the exact amount we are exposed inside, because of both elimination processes and secondary reactions, the latter which might leave even more insidious or harmful byproducts. Heat process products build up in tissues and cause telomere attrition, which is to say accelerate ageing on a genetic level, through diabetes and on its own. It was established that AGEs accumulate in organs, but that the rate of assimilation and build-up depended on the particular molecular species.
Predictably, AGEs bound to proteins, compared to free ones (with only a lysin moiety attached) are much more difficult to process and eliminate by our kidneys or intestinal bacteria leading to various chronic diseases, and so the rate of accumulation in our cells is much higher. Foods rich in protein and fat seem to have a higher content of CML and Methylglyoxal; in contrast, bread crust has lower content of these two AGEs. This echoes the fact that the danger of cooked meat far exceeds that of merely heated sugars which look a lot more like what the body is used to and can handle.
The blood-brain barrier is no more impermeable than the intestinal barrier. Both are merely custom checks points. So even though the brain makes use of dedicated systems (microglia, astrocytes, dendrocytes, perivascular macrophages and as we think, the misfolded proteins aggregates) it also use the standard immune system, in the form of tissue-resident T cells. Most if not all organs, have their own tissue-resident immune cells, maybe acting as liaison with the rest of the body with all things regeneration and immunity-related.
The term of “oxydative stress” is frequent in literature but improper, too reductive. The phenomenon does not limit itself to the production of chemical species called free radicals and other oxidizing species. Heat-altered species damage cells whenever the latter attempt to metabolize substances them: free radicals are most often the consequence, not the cause. They make us fat, regardless of actual calories or fat amounts. Abnormal molecules do the most damage when taking a structural role (bad fats and proteins), causing for example ingrown nails, tone and elasticity, severe skin aging, and partly varicose veins (poor smooth toneless muscles). The half-life of ROS’ half-life is very short: they do not accumulate with time, unlike abnormal proteic or fatty compounds in structural functions", increasing the background level of ROS and thus mutations, finally ending in either to tumors or necrosis as the rate of cellular turnover exceeds regenerative capabilities, repair mechanisms or the number of stem cells reaches zero.
Another argument against the toxicity of AGEs, is the fact that weight (a proxy for a higher consumption of food, thus AGE-rich junk food) correlates inversely with sanguineconcentration of AGEs but directly and strongly with fat levels. This proves a long-standing talking point of instinctotherapy: The body actively sequesters in excess fat molecules too dangerous to remain in circulation or be taken care of immediately. We stated the function of amyloid aggregates in sequestering AGEs for later disposal and it appears they also play a role in that capacity in fat cells. AGEs bind RAGEs in fat cells and accumulate there (secreting inflammatory adipokines, thus causing insulin resistance, though the role of the latter in “waste fat” metabolism is unknown.
White adipocytes (fat tissues not used for thermogenesis) reacts to exposition to molecular pollution by storing misfolded proteins in their lipid droplets. This directly confirms the observation made for decades already, that fat is a primary location for the body to sequester dangerous substances for later disposal, and not just a consequence of an excessive caloric intake. In that regard there appears to be two kinds of fat, (mostly) pure healthy fat which is easy to either dispose of or mobilize for energy, and denatured fat. It was observed, that, forcing the body to tap into that source of calories during too long a fasting or too restricted a caloric intake can be very dangerous as it also forces those molecules back into general circulation, which the body kept sequestrated for a good reason.
Commonly, this disturbs the nervous system (ramping up feedback levels) and muscular functions, from localized myoclonies to life-threatening heart issues, due to AGEs messing with mitochondrial functions (energy metabolism in cells).
Because increase in weight is directly proportional to the increase in those molecules, so while the kidneys do try to compensate by filtering more sugar out of the bloodstream—thus tiring their kidneys—they still can not avoid enhanced oxidative stress and microinflammation. Moreover, excess fat is a ticking time bomb, which lean people do not have to worry about. The moment this fat is put to use through exercising or whatever other physiological reason, the poisonous stock unloads. The density in molecules or the overall toxicity amount is not linear either, but the body seems to store the most dangerous in the deepest recesses: this explains why too much exercising risks opening up vaults of exceedingly toxic molecules along with the staggering frequency of cardiac arrests in professional athletes.
This is also the danger of excessive sudden fasting, which the mainstream is eager to ascribe to low fat levels (<6% BMI) in and of themselves. If push comes to shove the body will tap into its deepest reserves, including inside cells, regardless of their content: this is a normal and very beneficent regenerative process. Such storage habits are natural, however both the amount and danger levels of molecules not just rare but wholly unknown in nature (save for rare wildfires) that cooking brings, can exceed the body’s capability to handle the load through the usual elimination pathways.
The answer to discover our original food range, is to observe primates:
They feed only on natural food, neither mixed nor processed or seasoned
They are guided mainly by smell and only then by sight, color or memory.
Under optimal conditions the following pattern invariably applies:
attractive smell → salivation reflex = physiological need → luminous phase, an excellence in taste confirming the smell.
Saturation of need is indicated by an unpleasant sensation in the mouth: acidic, bitter, raspy, difficult swallowing etc, meaning that enzymes are missing.
Forcing us to stop eating, as continuing regardless causes a painful sensation in the stomach, if not immediately the mouth.
Some food have a stronger character than others for instance pineapple (that contain a protease) will make you bleed from the moment your body stops producing anti-proteases. This doesn’t mean pineapples are inherently better though, all wild food can become unimaginably repulsive to the point of throwing up.
The wideness of tastes they can take depending on your inner state, is a crucial characteristic of wild food compared to domesticated or selected species—which can’t be as rich in taste nor as harrowingly distasteful when the body refuses it. Just bland in comparison. It is a dialogue, an active exchange, like talking but in the primeval common language of chemistry and maybe more. Ultimately the level of delight we can attain, exceed by far anything cooking can invent: in fact it only ever tricks our taste buds, playing on a few strings discordantly while a original taste is a symphony in comparison.
(Wild) animals do not have our intellect, they spontaneously obey their senses to regulate the quantities. Too dumb to disobey nature. Influenced neither by dietetics nor the fear of missing, a good smell is followed by a good taste, they continue to eat, stop at the first sensation less pleasurable. It must be understood that metabolic needs evolve slowly therefore kept very close to those of our ape cousins.
Precise menu content:
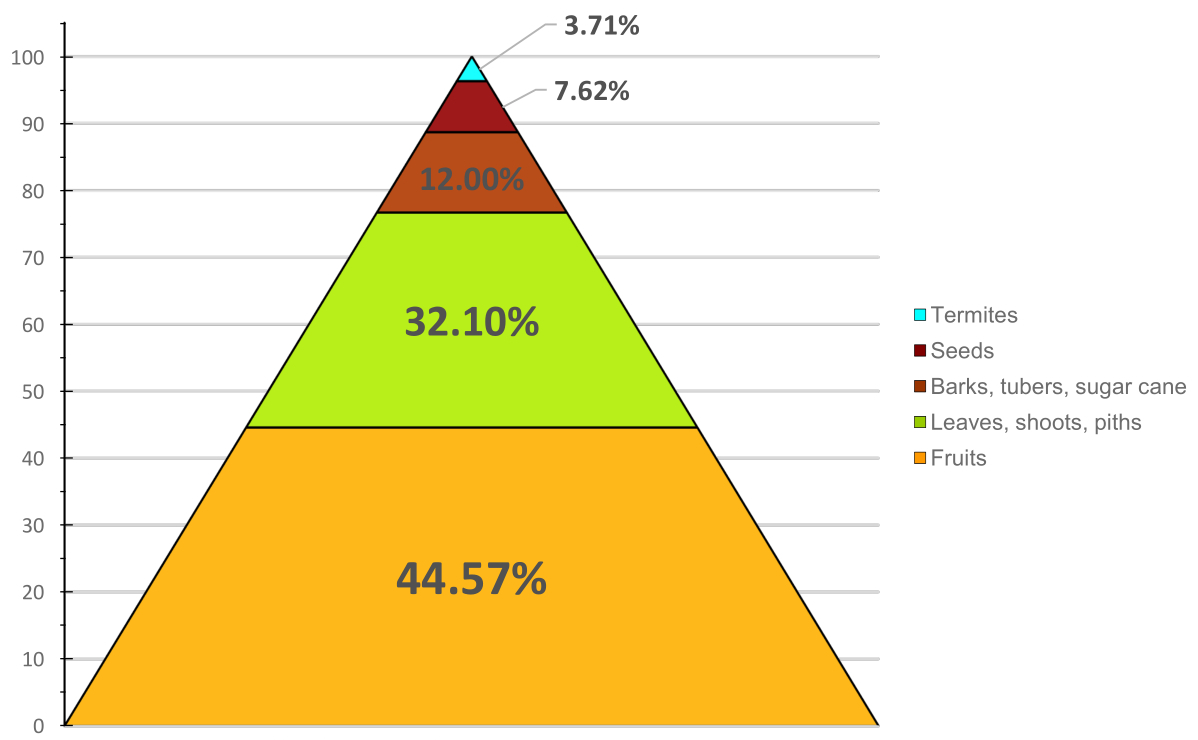
Fruits (pulp + seeds) > 50 % weight
Other plant foods: stem pith, leaves, petioles, seeds, flowers
Sometimes small mammals, up to young antelopes. Or smaller monkeys.
Honey, eggs, soil
Mushroom and termites, insect larvae
Bonobos
Bwindi Mountain Gorilla
Ngogo Eastern Chimpanzees
Rio Muni Central chimpanzees
Modern humans, USDA, 1992
We find that monkeys (like bears and many omnivorous animals) to be very opportunistic. They show clear preferences but the most seems to have varied food in 4 categories: carbohydrates (fruits, honey), vegetables (greeneries !), animal proteins, vegetable proteins. Depending on local availability, it is possible to rely on honey or a small variety of fruits… for a limited time.
No primate eating more than 5% of animal products overall. Even wild snow monkeys spend a large part of their day foraging, mostly fruits, but depending on what is available plants, small animals, insects, farm crops, and even soil, and a lot of bark in winter. With their intelligence, prehistoric men would have a much easier time finding the above which they prefered, while meat (a fortiori raw mammal meat which is the least appealing of all) is only ever a stop-gap measure for apes.
In practice, proteins constitute ~ 15 to 20 % of our intake in weight. Insects have revealed in fifty years of experience to become over time, for people practicing assiduously this diet, the most appreciated proteins to our gustative buds, much more than meat and eggs or fish.
Seafood (crustaceans and bivalves) are phylogenetically close to insects and thus therefore very far from us, while meat (especially mammalian) has molecular structures close to ours, therefore more likely to mislead the immune system, at least in the presence of excess quantities. More simply, even for a long-time practitioner with a relatively regenerated body and sense of taste, wild meat has as woefully or wonderfully strong a taste and stops you very quickly: That means we are not made to eat a lot of meat. The irreplaceability of insects is also understood by their easy access in nature, which makes our adaptation to them an evolutionary and logical necessity.
Walk through a virgin forest like the Białowieża Forest in Poland and you will see the ground littered with huge dead tree trunks, swarming with worms and insects. In a climate supposedly cold and inhospitable for a naked monkey ! So in a tropical climate, without human deforestation, abundance is a certainty and the primitive tribes of the New World know it well.
Larvae of barbels are found—in Europe!—inside oaks, beeches, ashes, willows, chestnut trees, apple trees, lime trees. Insect eating is not very developed in Europe (probably because of the omnipresence of dairy) for cultural reasons. It is commonly and falsely believed that humans can not digest chitinase, because many people do not present chitinase activity in their stomach. In reality we just don’t eat enough chitine, so we don’t produce chitinase, unlike people for which insects or crustaceans (seafood) is a regular part of their diet.
From South Asia, a whole industry could spread to the world
There is an element of intuitively danger to meat, which has been perceived all thoughout history, by Pythagoras and others1. Not to mention the ecological damage of hunting and farming and ethical problems of this industry. Observations indicate monkeys have a clear preference for eggs and invertebrates, followed by reptiles and birds.
An objective point of view would take into account the practices of monkeys, consuming meat in limited to non-negligible quantities. Total deprivation of animal proteins (which are not limited to meat) causes deficiencies, which explains why vegans invariably end up as thin as Auschwitz survivors. Monkeys are not carnivores like lions or dogs, jumping on the prey and gobbling it up alive, fresh. Exception of entrails and some organs, neither our dentition or digestive system are tailored to digest fresh meat.
Thereby primates are for the most part occasional, opportunistic meat eaters. So it makes sense that we evolved to consume bodies found in the wild, already dead and quickly rotting in a hot humid tropical climate, quickly emitting a strong, attractive scent. Even predators like dogs routinely instinctively their game in the ground. Henceforth, unsurprisingly all populations eating raw meat, like the Inuits, classical Romans and some people still in the 19th century, prefer aged or downright rotten meat.
The maturation or ripening usually is done by storing game in a cool place for a variable period of time, 8 days or more, in order to tenderize its flesh and obtain a particular flavor as germs in the intestine invade the tissues and break down the proteins. Mushrooms are also involved in the practice of “dry curing”, where they form a kind of hard green crust on the outside of the piece of meat, that help the enzymes and add flavor.
We no longer push aging to the point of “altering the scent” as Montagne advocated, or Brillat-Savarin, the inventor of the eponymous cheese which in his own words “bothered all his colleagues with the smell of the game he brought in his pockets to have it aged”. This would be almost impossible today, not without reason: Foreigners eating for the first time in developing countries often get “turista”, an explosive intestinal reaction to inferior food “hygiene” conditions. This is a way of saying that the intestinal flora has been disturbed and must take a few days to change and adapt.
Our advanced industrial populations live in almost total asepsis, compared to poor countries. It is no exaggeration to say that Westerners live on a permanent drip-feed of medicine, and without it infant mortality would immediately skyrocket and life expectancy drop. But experiences shows it takes only a few weeks to restore a proper biote.
Catching small game is not difficult either: Victor de l’Avéron, a wild child found at the age of 12 in 1785 by Dr. Itard, naked and unable to speak, insensitive to heat and cold, could spring like a gazelle, catch birds in flight as a cat would and pluck them in an instant.
But falling in the other opposite extreme, claiming we abandoned fruits because we can not compete with other animals in climbing is equally nonsense. True, apes have four opposable thumbs, their limbs’ proportion is much better suited to arboreal locomotion and allowing for efficient swinging. Yes, the evolution of our morphology shows a shift from an arboreal to a predominantly terrestrial lifestyle over the last three million years. But these reflections cannot determine how adept the original man was at moving through the trees. A good approximation can be reached however, by studying the ‘wild children’ who lived in the forest. Wild children are typically described as climbing trees to escape animals.
It has been said that the human ankle is fundamentally different, preventing it from climbing trees effectively as apes do… Yet some African tribes can still climb trees 20-50 meters high with no special skeletal adaptation, due to simple conditioning, albeit over a number of years giving their calf muscles’ much long fibers than those of farmers, and allowing their feet to bend up to 45° as opposed to the normal 15 to 20°. And we know for sure genetics has nothing to do with this, because their women do not climb trees, and their fibbers compare to those of non-climbing tribes. We are also still quite good at moving quickly from branches to branches (brachiation).
This means paleolithic humans were excellent at climbing and navigating trees by brachiation (swinging from one branch to another with the arms), and so could you, well enough to fetch fruits. we never quit the trees completely, or suffered the competition from other animals that would have pushed us in the Savannah to hunt big games as many claim. The same could apply for most so-called key differences between us highlighted every now and then.
The adaptations that have taken place all appear to be slow, multiple and gradual, and almost always present to varying degrees in our cousins for millions of years already. Fruits have always been—until now—the bulk of our food.
Behold Original beauty
We are still—without doubt—adapted to ripe fruit, and often birds are adapted to other fruits than us and only fall back on our agricultural products to the extent that we have greatly reduced the diversity around which these species have evolved. We have ruined nature as a whole, creating an image of scarcity and permanent shortage in a wilderness that is supposed to be abundant. This abundance and diversity can still be seen in the Kazakh forests, inhabited by omnivorous bears with similar tastes to ours. If our European woods show few edible fruit trees, it is primarily because we stopped caring for them and living in them tens of thousands of years ago, except for some Amazonian tribes.
In practice, we never wish to eat meat more than a few times per month, preferently fish or reptiles. Wild raw meat is no poison, but would not be consummed in nearly so big quantities, thus solving the moral issue raised by vegetarians. In absence of insects and fish, the body can definitely default to it for a significant amount of time if need be, however to conclude that the Ice Age made cooking a necessity is a fallacy: Many Ice Ages and Interglacials succeeded for the better part of the last million years. It stands to reason we adapted to them, and to a largely (raw) carnivorous regime… which systematically causes troubles of all kinds, including tumors if kept for too long.
Moreover, people used to move, a lot: why would a tribe stay at a place with nothing to eat but animals proving unsavory and annoying or dangerous to catch compared to fruits, weakening us the longer we keep on such an unbalanced diet ? Inuits, until recently total rawfoodists, age very fast, because they consume only meat. Any Animal would run and explore new territories more suited to it, why wouldn’t super-intelligent primates do the same ?
We definitely underestimate the amount of food a wild forest would provide, because we ceased to live in them and maintain our ecological role a long time ago. Ending an obligate co-dependent symbiosis when we started cooking, dense forests cease to be primate forests anymore. In the span of a few millennia, without dissemination fruits either disappear or degenerate into unadapted forms. The process finalized when we drove away or genocided bears, the only comparable fully omnivorous animal with an active metabolism.
Paleoclimatic modelizations are erroneous when depicting the Ice Age as a barren, sterile no-man’s land frozen half of the year. Ice ages are defined by winter cold enough compared to hotter seasons, so that ice does not thaw entirely, but either sustain itself or pile-up over the years. A good part of Swedes, in particular the edges of hills and mountains, are in that situation, and forests do not shy from nearing glaciers. The same occurs in Switzerland, owing to its topology. It is a matter of temperature averages, and most of Sweden isn’t barren half the year, but features summers hot enough to get naked comfortably.
The part of Europe not under glaciers knew climates varying between modern day Sweden, Switzerland or Norway, with inland masses (and coastal areas, depending microclimates) had a lot more of the typical continental climate (unlike the current temperate one), with strong winters and warm summers as if suddenly placing France into Ukraine or Poland, or Russia for more Northern places. One could hardly call Ukraine (at worse, Finland) regions deserts. Instead of the lack of fruits or impossibility to grow them, the development of hunting, might find another explanation in addiction to cooked meat.
People relying on meat quickly lose the ability to eat a lot of plant-based food, and their population correlates with animal population density even when the seaons should allow a plant-based diet, thus leading to a very inefficient and irrational use of the available resource.
Bodies need no intellectual guidance from dietetics for the good reason that it knows its own needs to the second and to the milligram of substances: Many people ignore or would deny the possibility for their nose to indicate anything, or that food’s scents can change from one moment to the next. They just never experienced it. It is to realize we have been subjected to cooking since birth. Every instinct requires a concomitant learning ensuring its correct application. Herring gulls, for example, drop hard prey from heights onto hard ground, to eat the inside, crabs for instance. Young birds up to one year old tend to fail ridiculously: catching in the air what they just dropped, dropping it on water or soft surfaces or do not fly high enough and must repeat. This contributes to the high mortality of this species’ younglings.
These feeding instinct behaviors require extremely complex cerebral motor coordination, flexibility involving an ability to learn various behavior patterns depending on prey, soil, wind, competitors. Youngsters have the instinct to try but understanding the relationship between hard surfaces, height and success takes time and often fails. Sometimes they lose interest in their prey, which is then stolen by a competitor.
In some species, such basic behavior requires a learning process. The same is true for feeding: the instincts are actualized in the key moments of childhood. This notion highlighted by Konrad Lorenz is fundamental for food and love. Thus the very young baby in front of raw food, will immediately have the innate reflex to smell, to open its mouth if the odor is appropriate, and will turn away its head if the odor is not very engaging or even will spit it out. Many parents will attest to this. Within a few weeks, most of them show an interest in meat, and chew to extract the juice because they have no teeth.
On the contrary, in the culinary context, an habituation born from the constraint of denatured, industrial or otherwise processed foods, compromises the baby’s ability to assimilate the scale of tastes and smells in relation to inner needs. Cooked flavors change very little, because the genetic programming of senses supposes the contact of unadulterated raw food. Obedience to parents and conditioning through the vintage command finish your plate eventually replace the baby’s instincts with eating habits, dietary beliefs and a permanent inner metabolic disorder.
The child born raw (or wild animals) on the other hand gets to know his body and its subtle signals by associating pleasure to obeying natural laws and discomfort to disobeying them. Acquiring this natural acuity of smell and proprioception during adulthood is difficult, it requires a lot of discipline, and the application of some rules developed with the years regarding the number of meals, their quantities, some hints of satisfaction or overload, etc. These are all crutches, but absolutely essential ones for all those who hail from a cooked society.
Maternal diet from the moment of conception to the delivery is known for certain now, to impact the child’s health for its whole life. Even disregarding abnormal heat-related molecules, cooked food’s level of vitamins can be tens of times lower than in raw food So by indulging in cooked food your children are robbed of a normal development, given deficiencies which may potentially not be corrected completely if said child is given raw food from day 1.
In “primitive” societies, juvenile humans (preadolescent) can largely fend for themselves, deal with the danger of predators, diseases, and provide some of their own food. In some of them, children are left on their own device immediately after weaning, and spend many years until adulthood living with other children in their own parallel society.
Since cooking and later the Neolithic Revolution with the advent of agriculture cerebral disturbances have increased steadily, warping our very psychic functioning further and further from its natural state, which all unstressed wild animals share.
On the opposite cooking induces a neurologic feedback loop to repeat mental patterns, constantly feeding cerebral centers the product of their own processing time and time again and drifting further and further from the original signal, be it a physical stimulus or the first impression. Like a micro and speaker in the Larsen effect. Thus, this automatism mental, we call feedback, as if the brain actively feeds on its own noise. The consequences on sexuality, described elsewhere, are immeasurable.
Nervosity reduces drastically, making normal people look like advanced mental cases in comparison, without exagerating. Glutens in particular (group of proteins specific to true cereals from the Poaceae family including wheat) is demonstrated in clinical studies to significantly increase symptoms in schizophrenics with a link to coaeliac disease and autoimmunity as well. Sweetened drinks (soda, chocolate with sugar, coffee with sugars, anything with artificial sugars really) have been linked several times with dementia, brain aging, onsets of Alzeihmer and smaller brain volumes.
The level of calm one can obtain just from rawfood is hard to fathom save for seasoned meditators. One really has to live it for oneself: the number of situation one feels normally overwhelmed by impressions and emotions in daily life and acting on them or thinking it’s a big deal is staggering, it is a new way to go about life, that alone offers nearly for free a peaceful state of mind worth a lifetime of investment in spirituality to most of the time, little to no avail.
We discover these uncontrollable urges are but dysfunctions of the brain excited by the constant influx of denatured molecules, now acknowledged in their psychogenic effects.
Because of this constant mind poisoning since early babyhood, the brain actually wires in a pathological paranoiac (and dissociative) mode that only a long exposure to a balanced nervous state can correct, although imperfectly. Hence the continued importance of some mental discipline, to recalibrate ourselves along natural lines of behavior and feelings.
Animals on the other hand, do not have our in-built levels of self-control or metacognition, turning them from gentle to violently insane in an hour: cockerels and pigs fed with (heated) grains and leftovers attack their peers or females, leading sometimes to an orgy of violence and blood. The problem is well-known though the association with dietary conditions, hardly so. In the same vein, Africans have always been raping and eating one another since time immemorial.
Wild animals (or domestic animals not fed with processed food) act much more harmoniously, tolerating each other and very rarely indulge in fights leading to injuries. Dogs stop barking ceaselessly at the slightest leaf falling, cats stop hissing and fighting in the middle of copulating. The list goes on and on.
As our mind calms, many subconscious mental blockades whether born from habits or childhood conditioning dissipates, allowing for a richer inner life more in touch with deeper meanings in life and relationships. Cerebral functions improve and the constant fog that followed us all our life evaporates, making school work easier. Compulsive or obsessional ideas lose their strength, progressively discarded. Things we were taught as evil or diabolical, appear not so bad after all. Negative feelings, shock and trauma, become much easier to deal with, and fade faster.
Cooking on a small brain
Sex becomes an entirely different affair. Men loose their compulsive need for gratification, which enlarges into a more satisfying experience, and nourishes higher communication between partners and a keener perception of each other’s wants and desires. No more uncontrollable erections, no more muh dik: they only rise when the moment calls for them. Men stop seeing women as walking pieces of meat.
The tolerance for mental and physical exertion skyrockets and sleep improves. Fatigue can stack up a lot more, for a day or more if need be, without stimiulants, until of course has to clear up one’s debt.
The rejection of the germ theory in favor of a new understanding of viral phenomena as symbiotic and beneficial for our health frees the mind and increases the capacity for regeneration, allowing us to live fully without a care in the world. The need for nearly any medicine, save for a few cases of surgery (dentistry and accidents), is no more. In good conditions bruises and hurts, even serious, not only do not hurt at all except on the very moment of injury, but heal up several times faster than a normal person without any disinfectants, barely any care even letting dirt and whatever on the surface, just for the lulz. We can develop an almost superhuman healing factor.
Blood clotting takes less than a minute at least for life-critical outflows. It appeared that the considered normal levels of pain is not natural but caused by a constant high level of autoinflammation, raging on the smallest occasion. That explains why wounds just do not hurt when let them still and the work to do its work. The body can regenerate damaged internal organs to the extent to cure almost all diseases, and slow down the degenerative part of aging to a significant degree. We age much better, and slowly, even without training. Samewise, the amount of flesh that is possible to regrow when still enclosed by sufficient surrounding tissues approaches the healing factor of the likes of lions, healing big wounds healed in no time nor a care in the world.
Years showed the complete eradication of infectious diseases and cancers alike when the latter are not too advanced. Even then, it allows for a painless death with full mental faculties and no medication. Similarly, those born or at least raised on instinctonutrition will never worry anymore about tooth decay. Wild animals with a balance diet do not develop caries, and it wasn’t uncommon for people before WW2 to never have a single cavity in their entire life.
The body is much more resistant to training as well, and recover faster, but even more surprisingly training had become unnecessary to keep in shape. Animals are basically born with rippling muscles and maintain them regardless of training, as long as they have good quality food: May they slacken as they please, they do not fatten. Men on the other hand, are known to lose muscles without constant exercising. This comes both from the impossibility to substitute our intellect to the precision of our body’s dietary instincts, and from the low-quality proteins constantly breaking down in our muscles.
Aging slowing down, we keep a slim/athletic physique as long as we keep a varied enough diet to continue on the cleansing process, and most of all if we keep clear from excessive animal meat. Excess proteins do the same as unnatural proteins, and shorten lifespan, eating away at our healing factor. The best part of our lifeforce is constantly undermined and spent on fighting off the autoimmunity caused by cooked molecules.
In this section I comment on the only study of raw food practionners (for any definition of “raw”) lasting reasonably long with a significant sample size. We wish to demonstrate that despite the obvious and expected biases the analysis demonstrates, the effects they conclude to be detrimental, once recontextualized in the paradigm of instinctive rawpaleodiet, are consistent with our own observations and the theory, except that they are neither detrimental nor pathological.
Conclusions: The consumption of a raw food diet is associated with a high loss of body weight. Since many raw food dieters exhibited underweight and amenorrhea, a very strict raw food diet can- not be recommended on a long-term basis.
The first issue was to lump together participants in excessively broad categories, with omissions testifying of the complete ignorance of the researchers, about nutritional needs or an ideal primate diet. But this is a fundamental methodological issue: we can not make sense of such unstructured data as interviews without a preliminary understanding of the underlying reality. Only 44% of the sample eats meat with or without fish, 32% only keep eggs and milk, and the rest is vegan. So there is a massive bias against meat, and no distinction made between wild animals or livestock. No mention made either of seashells (marine products beside fish), or God forbids insects, despite both being major protein sources, very distinct from meat. So any conclusion about weight effect should be taken with a grain of salt by default.
Other biases include:
No notion of bodily composition, the difference between muscles and fat (and between good and bad dat) is completely discarded. Someone having before starting 5-9 kg of excess fat but lying in the normal range around 20 BMI, will loose his fat and reach and fall in the “underweight” category. Yet he would not lost anything useful.
This “100%” category include both meat eaters, vegans and fruitarians, leading to the introduction of chronically starved people influencing the average disproportionately. If loosing is weight, recreating healthy tissue on the other hand requires a good balance and exposition to varied natural proteins (eggs , seafood, wild meat, insects).
What raw food people do is usually very far from instinctonutrition, from a natural, animalistic alimentation: in French it deserved the name “crusine”, which is to say they do everything that comes with cooking (seasoning, mixing, sprinkling), just without actual cooking. They also often eat products not even raw in that limited sense (cold-pressed olive oil, “raw” oilseeds dried in the sun at more than 45°C). This can lead to very unbalanced, even dangerous diets.
Yet despite all this, their conclusions are laughable and contradicted by their own, actually very positive data: Nearly all participants who ate at least 90% raw lied in the range 20.2–20.6 kg/m². BMI was inferior to the norm (< 18,5 kg/m²) in 14.7% of men and 25.0% of women. Despite the initial loss, weight always stabilize to a functional level. What people always loose, is nearly always useless fat.
We see also a fantastic confirmation of the pathological nature of menstruations as experienced by humans:
About 70% of female participants noticed a change in menstruation since changing their diet.
Among women of childbearing age, 23% suffered from a complete absence of menstruation. Women with partial to complete amenorrhea ate more raw foods and had a lower BMI than other women.
This is excellent: Menstruation disappears instinctively but it happened to women on instincto women too, with no issue on fertility. The people who lost the most weight compared to before, in fact lost their excess fat and the denatured molecules that it sequestered. Periods in fact have no causal relationship with fertility, because their real function is to serve as an additional elimination pathway (doctors of the last century justly spoke of periods flowing through the nose).
Hence women whose weight stabilized, swapping bad fat for muscle no longer requires periods, while reintroducing cooked or raw meat in excessive quantities amplifies the outflow.
Cooking makes us all too fat, we are the fattest primates, with the highest BMI, despite a metabolism seemingly 27% higher than chimpanzees. Women in particular are all too fat and physically pathetic. Like for any animal, they should be on average more agile than men of the same size, lighter on their feet, and packing the the same power for the same body mass. The idea of needing extra fat to bear children is ridiculous cope. Nor are we bears hibernating and building for the winter season: instead we use our brain to find food or make reserves… like squirrels with brains the size of a walnut.
We make the case that fire was never needed to survive but instead caused of all bodily pathologies and suffering, directly or indirectly. Because we attack something so close to people’s heart, maybe dearer than anything, anyone or any belief, something most think we can not even survive without let alone thrive, a logical reaction would be to wonder, why then it started in the first place ? And how did it become universal ? Men have used and controlled fire fully or partially for more than a million years, but we believe that for the most part, it was not associated with cooked food, which came only recently (compared to the age of our species, which we will come back to later).
In 1995, the British primatologist Richard Wrangham formalized in Catching Fire what everyone in paleontology and anthropology, not to mention the general public, had thought for a long time, namely the hypothesis that the reduction of the jaw and the size of the teeth accompanying the growth of the brain in Homo Erectus and the species that followed it would not have been possible without an early mastery of fire allowing the cooking of food. Thus cooking, by outsourcing part of our digestion, would have allowed us to waste less time and energy eating, devoting these resources to developing our brain and properly human symbolic activities.
Wrangham is the quintessence embodiment of everything rotten in modern science: Not only is he an enemy regards to cooking but he spearheaded the notion of Chimpanzees being raping killing machines viciously waiting for the smallest overtures to gang up on females and children.
Moreover, his thesis would date the mastery of fire to nearly two million years ago, whereas the certain habitual use of fire in households dates, for most specialists, to 500,000 years at most… as reported for the Qesem cave which associates the earliest control of fire mainly with the first Homo sapiens and Neandertals. In 2012, microstratigraphic evidence (based on microscopic analysis of sedimentary layers, looking for chemical or isotopic traces) of the earliest fire use by hominids in situ one million years ago in the Acheulean strata of Wonderwerk Cave, Northern Cape Province, South Africa, arrived. This is considered unequivocal evidence for the habitual use of fire by Homo Erectus.
However, Wonderwerk cave does not present the telltale sign of mastery of concentration of burnt remains in hearths. After, we see an absence of evidence until ca. 250 ka, proving fire was never necessary to survive in Europe’s seasonal sub-zero temperatures, including in Northern regions or England. Our lineage has been living here for 2 000 000 years straight: we adapted, either living naked or making occasional use of animal hides.
Earlier claims from sites like Schöningen, Terra Amata, and Vértesszőlős have faced substantial criticism regarding either dating accuracy or the anthropogenic nature of the fire evidence. So the most secure and convincing sites for early controlled fire use in Europe are Valdocarros II (Spain, ~250ky), Menez-Dregan (France, ~200ky), and Bolomor Cave (Spain, ~200ky). All show features indicating of deliberate, concentrated and repeated fire use.
Posterior or overlapping those dates, several European Paleolithic sites lack secure or regular evidence of fire use often despite intensive human activity: Gran Dolina (-450-200ky), Galería (Atapuerca, -350-200ky), Arago Cave (-350ky-200ky), Pech de l’Azé IV (-180-50ky). These last two sites altern fire-rich and fire-poor layers, both occupied for thousands of years, suggesting that fire use was either irregular or absent for prolonged periods.
So all in all, we can conclude, at minima, that cooking, or fire at all, could not have been a frequent occurence before 200ky ago, nor as ubiquitous as it is now before Cro Magnon circa -45ky BP. And if we consider secure hearth sites, it is more or less to prove the cooking of food before eating, as opposed to peole just nurturing fire with food remains of the last meal. This only illustrates well how the overwhelming majority of specialists (and common folk) just can not mentally separate fire from cooking.
The mainstream hypotheses come down to: eating more meat, scaring animals at night to sleep in peace (especially since we no longer sleep in trees), lighting, keeping warm. We believe that in nature, fire has never been more than an accessory for our ancestors, whose body and brain were perfectly adapted to not only survive but thrive. None of the listed reasons really hold up under scrutiny. Our natural abilities ensured our survival in all environments).
Firstly, as we discussed, meat becomes edible when aged. We do not usually eat huge quantities because it rarely meet our nutritional needs, not due to a lack of opportunities or intrinsic difficulty to digest without cooking.
. Most people, including scientists, think fire instinctively scares all animals… This is wrong. Lions or hyenas are cautious but not fearful, and you will lack a few toes if you think it will protect you at night. The same goes for wolves and bears. Moreover some are attracted by the smell especially as they learn to associate it with our food. We are not natural preys for any animal. Neither sharks nor tigers will instinctively pounce on us unless starving… or drugged. Besides numbers alone along with spears, deter predators interested in your children or pets, more than fire could. The common picture of weak prehistorical men only surviving the assault of cruel wild beasts with the product of his brain is a self-comforting myth for modern people.
As for lighting, of course, we are not cats, we do not have a tapetum lucidum which reflects light behind their pupils. Incidentally they have 150 million rods instead of our 120. But in the end, it’s not a question of being able to hunt in the forest as if it were daylight… but simply of managing to navigate through obstacles two or three meters around you, in the dark. And this is easy enough if the moon is beautiful, and the stars numerous… Which was always the case before the industrial age. There is a special training for night vision (scotopic), well studied during the Second World War. Simiarly “wild children” are often mentioned to have excellent eyesight, a fact corroborated by the active night life of apes. We obviously evolved to find our way around at night even in the darkness of rainforest forests. Caves are different case, but then charred bones, which become fluorescent, can be used to great effect.
Lastly, men like Wim Hof prove that we could easily sustain prolonged exposure to sub-zero temperature with no issue, running an Arctic half-marathon (21 km), bearing an average of -20°C for five hours. He also spent 6 minutes 20 seconds under the polar ice, 72 minutes in a translucent container filled with ice, and climbed the Everest with minimal technical equipment, in shorts, gloves and a cap only stopping at 7,400m due to an old foot injury from the previous year.
Meet Wim Hof or Iceman
And many monks develop similar abilities in Tibet through concentration and training, which is now being studied in the laboratory under controlled conditions. Difficult to guess what use such adaptability might have had for African apes: what sense is there to survive in places where nothing grows anyway ? We can safely assume that at no time did we need fire as long as they stayed where there was food to eat, which in any case excludes the Arctic Circle. Even so, clothing (animal hides) would provide more than adequate protection.
If charred bones and most burnt food remains (including a number of wild cereals) will always stay unconclusive, the same can not be argued for many tuberous vegetables such as manioc are absolutely impossible to eat raw, as are grasses (some cereals such as maize can though). Their place in today’s diet is explained by the opiates they contain, which make for a terribly addictive effect.
The first attested granaries date back to 11,000 BC. The consumption of cereals may of course be older. I am of the opinion that cooking must have appeared several times in several places, more or less accidentally, because consequent control and taking thousands of years to develop, allowed sufficient food security to concentrate populations in sedentary centers, contrary to what has always been the case. This article may point to an early, if short-lived, establishment of cooking and cereals, whose remains were found in Mozambique on stone tools associated with grinding stones and pestles dating back 100,000 years. But I would rather wait a few years because it seems early, and if it is indeed about cooking and cereals—which I am not sure of – as obviously not persisted. There might have been a technical use of these products, perhaps an early onset of an industrial process rather than a dietary one.
It is conceivable cooking could have fascinated relatively primitive populations in terms of intelligence, once a certain brain size threshold was reached, say, 800 cm³ perhaps, but then groups falling prey to this lack of instinctive programming against the product of our own intelligence, would become ill quickly and die from diseases, predators or be supplanted by healthier human groups. Indeed all animal breeders or farmers know the universal addicting effect that light cooking has on animals. But controlled cooking is not natural, and very difficult to pull off by chance as opposed to the much more frequent occurrence of unappealing charred remains in the trail of wildfires. So it makes sense that early attempts to replicate accidental “fixes” were probably immediately toxic, and were refined over time not to kill them too soon. And the “safer” the recipees, th faster the disorder and easier the addiction could have spred, and those groups with deregulated sexual instincts finally outbred all others once population could grow faster than diseases killed people.
But… this theory implies pre-cooking prehistoric humans were unintelligent, ape-like creatures, which is to say, Africans. The morphology of prehistoric Eurasians teach us the opposite: they were much smarter men than us. So while the addiction and alteration of behavior explain the diffusion of cooking after it had gained significant momentum, and and explain why reverting to raw food has been impossible since then. But this seductive view of the start of cooking as a chance drug addiction unfortunately taking off, can not be taken seriously anymore.
The origin of cooking is still a mystery.
The nature of human reproduction are all glaring symptoms of our present state being unnatural, unharmonious, either suffering from cooking and/or having degenerated from an earlier more perfect one, since at least the neolithic revolution. More precisely our reproductive function and tendency has been grossly exaggerated, overclocked, on a physiological level, breaking havoc in our health and mental balance, scarring our culture for millenia perhaps more than all other issues combined.
We should not have so many children, our biological cycle should be based on their needs. Cooking participates in a singular acceleration of our rhythm of life, for several reasons:
The molecular chaos represented by AGEs and other abnormal substances constitutes cellular stress due to their direct or indirect cytotoxic, genotoxic, mutagenic, carcinogenic effects, etc…
By directly amplifying the impulses linked to the instinctive reproductive program which explains the preponderance of heterosexuality cooking is accompanied by a profound change in the state of mind, and from there imagining a hormonal action induced on the gonads does not require a considerable leap of faith.
Compare this with the biological rythm of women with a diet rich in fat and/or processed food, in a traditional demographic regime, with constant intercourse and no contraceptive:
sexual maturity 8–13
periodicity of ovulations: 21-35 days, 28 on average
first birth (with constant intercourse and in absence of contraceptive): 8-13 immediate
Maximum lifespan: 85 (in good health), 100+ otherwise
Operational menopause: 38.7–60.0, with an average of 49.6
The normalcy of at least four constants so far must be debated:
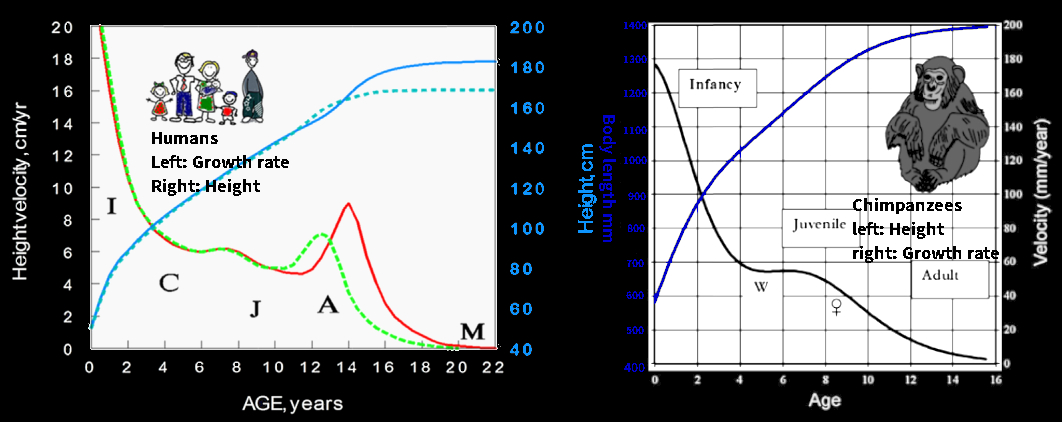
Starting from the widely acknowledged human growth curve anomaly among all animals: Usually we can see the height curve is rather linear during childhood, to show a weight of inflection and in adolescence, what is called the puberty peak: this does not happen in raw children, there is no puberty crisis, growth (and the development of secondary sexual signs) is slow and prolonged, with no identifiable beginning or end. On average they are a little smaller but this is not comparable to a lack of growth because they are stronger and much healthier: in truth we are too fat and of poor bone constitution, because of dairy products.
Human growth curve vs chimpanzees
Human Growth and Development in Basics in Human Evolution, 2015
We then see that although growth (and weight gain) accelerates during puberty, the transition from puberty to fully adult age is abnormally rapid. The onset of secondary sexual signs should be slow and continuous, instead of sudden and quasi-traumatic, as the majority of adolescents can confirm. Size does not matter, all mammals, including cows and mice, show a similar continuous growth curve, just more or less elongated, with little to no point of inflection, nor can we measure in them our strong hormonal surge associated with a multitude of health issues.
The juvenile phase exists in all intelligent social animals, which makes sense given its obvious learning function. This is the most dangerous period regarding post-neonatal mortality (predators and the danger of competing males, in the case of lions). This period is in absolute and relative terms the longest by far among the human species… With here significant differences between Africans and Asians/Europeans.
Our view is that there is a general, underlying factor in obesity and early maturation: molecular pollution in all its aspects, that it causes obesity. Guyclaude’s children reached maturity only toward 25, while being consistently more muscular than all other kids their age.
Little or no breastfeeding also relates to faster development. This can be explained in several ways: lack of breast milk leads to paradoxical development, directly stressing the body, endocrine balance, etc. Denatured molecules accumulate more because milk allows the body to learn how to better clean/get rid of them.
The lack of human milk also certainly leads to a paradoxical development, directly stressing the body, endocrine balance, etc, as we know its importance for the development of the brain. The Quechua Indians seem to feature by far the most delayed growth ever, ending at no earlier than 22, while still congruent with an admirably strong, sturdy body.
The classic undernutrition or apoxy explanation is inadmissible because their strength and overall health put city people to shame. Beside, Tibetans contradicts it, with periods closer in time to ours (16) but an ever earlier menopause (45 versus 52 here), as well as their overall aging. But unlike Peruvians, they eat much more (cooked) meat and worse cereals too. Recent studies corroborated that association. The fatter girls are, the earlier their first period..
These accelerations in growth (inflection point) are always so small relatively (regardless of size, in mice or cows) that they cannot be compared to ours, because it should be enough for them to look to get a clear picture. Yet instead we resort to dishonest mathematical tools to level the difference, erase our uniqueness among mammals. The observable difference between observations and the no inflection hypothesis, null hypothesis is so small (less than a centimeter, and often only a few millimeters) as to warrant exponential regression formula while simple plots suffice for humans. That process multiplies deviations by a factor of three… yet the result was minimal.
The other line of arguments claims that hormones and chemical biomarkers do support the notion of a transition to puberty. But no one denied this, for genitals growing must certainly start at some point. The point of contention is about the speed at which it goes, and whether it shows with the naked eye. Collagen and osteocalcin levels (indicators of bone turn-over) nor that of creatine and testosterone do not mean much if anything in practice if we can not see the result in terms of actual growth. One must consider the sensibility to these hormones, which appears to involve two to four times greater for the human arm or leg, and more than 10 times greater for human sitting height.
Most recently a new article claimed to end the debate by redefining the adolescent growth spurt altogether, arguing that the correct way to plot height against age requires raising height to the power of 2.5 first. Why such a gross amplification (timidly called “rescaling”) should be legit when the spurt is not only visible for humans with mere linear plotting but often drastic ?
The juvenile phase exists in all intelligent social animals, which makes sense given its obvious learning function. This is the most dangerous period for postneonatal mortality. Incidentally, note here significant differences between Africans and Asians/Europeans. There are two basic ways to get from infancy to adulthood. One is to develop quickly and minimize the duration of the nonreproductive period between weaning and sexual maturation. The other strategy is to develop toward adulthood slowly, increasing the time for learning and producing higher quality adults.
What we call “traditional demographic regime” (free sexual activity and no contraceptives nor modern medicine) is described as “high stationary”, because understandably Such parameters give rise to a very high birth rate, 4 to 8 deliveries per woman in the Middle Age but only because of the custom of marrying late in your 20s, several years past sexual maturity or even the fertility peak, and other regulatory practices. Backward African countries bent on maximizing natality show us the expected result in nature with constant intercourse, as per our instincts, which is an excess of 7 children, up to 12, without multiple births.
The only reason this regime is called “high stationary”, is because this high nativity is compensated by an equally high infant mortality, plus wars and famines: all factors leaving no traces before the Neolithic revolution, and that could not have existed before cooking. That means that in absence of social regulations of matrimony, a people whose food is somewhat rawer, somewhat more vital and richer in vitamins, thus enjoying a good health and no epidemics, around the level of the least disturbed chimpanzees would witness an explosive demographic growth.
A hard-boiled belief in the scientific community is that all species would tend to produce as many offspring as possible given available resources, and would fill the environment if predators or other environmental constraints disappeared. Some species require predation to maintain the ecological balance, on which many other animals or plants depend, including those they feed on. The point of life is to survive, not to increase beyond measure. And an unlimited multiplication would lead to the destruction of the environment, as we realize this bitterly today.
Without diseases and infantile mortality to compensate, a default natality of more than 3 children on average suffices to cause a net demographic growth, and be ecologically unsustainable.
Species like ours need to measure and curtail its reproduction to fit the bearing capacity of its environment, as shown by meerkats and wolves for instance… and chimpanzees. Despite no ape being a seasonal breeder properly speaking (we are fertile all year long) in both chimpanzees and hunter-gatherer populations, birth peaks are known to coincide with periods of food abundance, from season to season and year to year. Apex animals all develop self-regulating behaviors and since animals do not take long-term “rational decisions” we are obviously talking of unconscious mechanisms.
This monstrous fertility defies all logic.
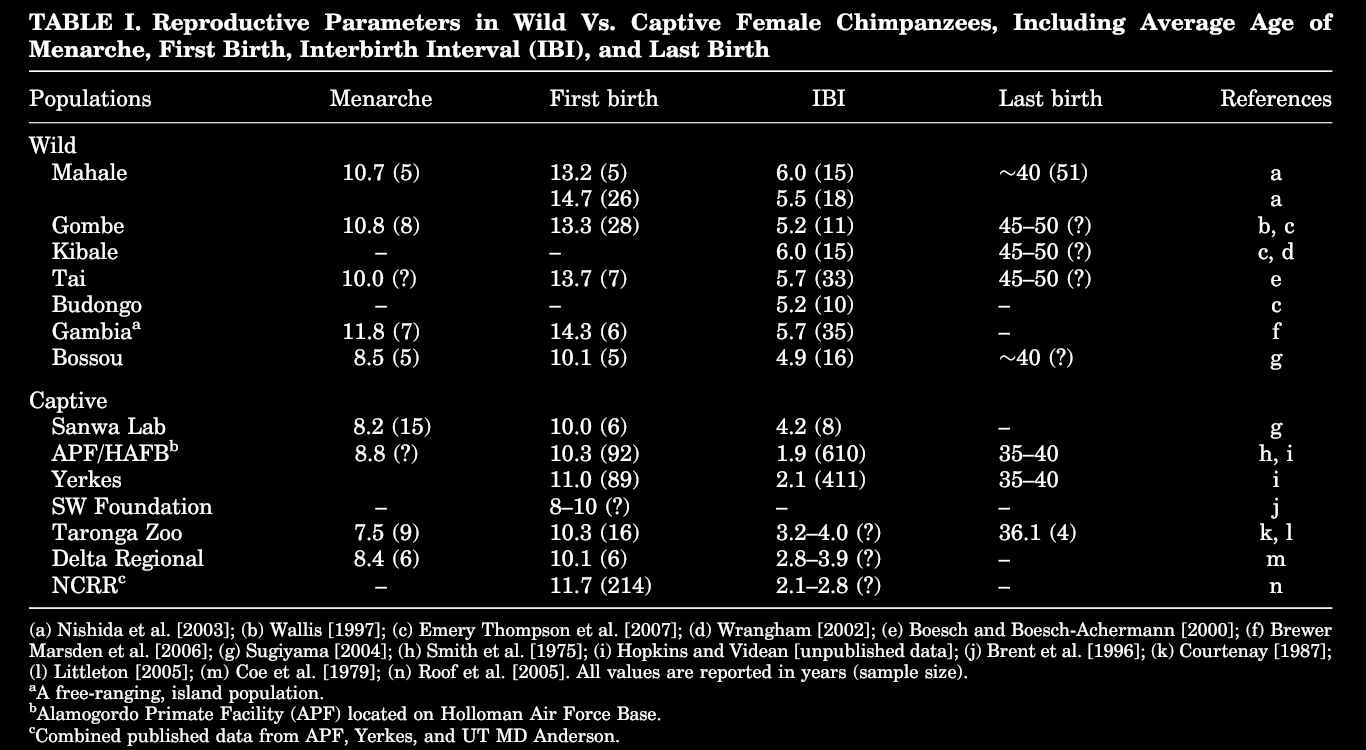
Let us look at the weaning/interbirth time variable:
Because chimpanzees basically mate constantly (like we do), despite a dominance of lesbian relationships most females are either pregnant, or with a child. So what delays one pregnancy from the other, is only the time taken for the offspring to wean.
How to calculate this for humans ?
In the US a fourth of all babies start out with baby formula, and half are given it by the 6th month, so people hardly ever breastfeed more than a year, against an average of 5 kids in wild chimpanzees. But it wouldn’t matter because today, with a “rich” diet breastfeeding only acts as a contraceptive for the first six month. And it is said that the LAM (breastfeeding) method only works if milk is the only food but this is not the case for chimpanzees, as from experience in just a few days infants smell and eat all kinds of food including meat, simply squeezing juices out of it for lack of chewing instruments.
The replacement rate for great apes is low, around 5, because of infant mortality (and taking into account the rare accidents) but in humans experience showed this reduced to zero. So in our case, more than 3 would be an ecological catastrophe. Yet even if the inter-birth time had not dwindled and we had to wait an average 5.6 years between births, so considering a fertility window of 30 years this would still leave room for a minimum of 5 children. With our diet menstruations disappear entirely in a few months without any adverse effect on fertility, as well as the famous pain of childbirth, after a year. The strong tendency for inflammation or autoimmunity caused by a permanent influx of denatured cooked molecules also disappears, releasing a treasure of bodily capabilities and resistance to muscle and psychological stress of all kinds. With time (more or less years depending on the choice of food) the tissues rebuild themselves on a stronger and more durable healthy basis.
Lilith
It is well known that captive animals develop significantly faster, breed faster and have shorter times of lactation per offspring, and shorter ovulation cycles. As a consequence their ovarian reserve deplete faster—reaching reproductive senescence early—and also age faster overall: every life history parameters reflect a shorter, faster life cycle. We can almost talk of a transition from a K to a r breeding strategy.
Compare this to the new human condition under instinctive diet:
adolescent growth spurt disappear, development is continuous, in height and secondary sexual characteristics
Periods disappear after one year of correct practice.
sexual maturity is not visibly before 18 accomplished, or older.
Long periods (in excess of a year) with daily unprotected intercourses may not lead to pregnancy, while in other circumstances, it occurs whenever women desired it, as if on command. Precisely what the Trobriandese people claimed.
Historically in regions less dependent on typically neolithic-related “advancements” (less milk and cook meat) menstruations used to start closer to the age of seventeen than to thirteen, and in some regions after 202. Several doctors attribute this delay in puberty to the arduous work to which girls from the countryside are subjected. They are aware of the city/country differences and grant a certain influence to the “passions” which agitate city dwellers earlier. It is the climate, finally affirms the doctor from Bourg‑Saint‑Andéol, which explains the advance in puberty of the girls of this small town. The climate hypothesis dominated the discourse on puberty for a long time.
There is a certain astonishment among these doctors: ultimately, the late age of menarche seems to them to be an anomaly, pointing to a poor economic and health situation. However, some are reassured, this late puberty does not prevent high fertility; in some cases, the late age at first menstruation can be compensated by also late menopause, which extends the period of fertility.
“Pale colors” are often associated with those girls in the moment of their period. But since these children are nevertheless always strong and healthy, we can assume that this “delay” in constitution does not correspond to a lack of development, but to a natural state. It is not malnutrition. On the contrary, these observations show that physical strength, the strength of an adult, is unrelated to maturing as an adult. In a sense children used to grow tall, but not old, or very, very slowly. Even then, the label of malnutrition (and being “underweight” often sticks to what are simply children growing at a more natural rate, or not as fat as nearly everyone is today. And could we deem “pathological” a reduced fertility, when the norm is absolutely unsustainable from an ecological standpoint ?
So, late periods correlate with a less abundant onslaught of denatured molecules, and idle children from the city age faster and weaker. This comes from the accumulation of denatured molecules, in the same way that puberty conditions the appearance of pimples etc: a growth which occurs stops seeing the toxins concentrate and require additional outlets. Additionally an urban setting offers more abundant and more processed food, meat in particular. While Republicans became obsessed with making women breed more and sooner, other physicists had more common sense, understanding that later menarche is a sign of a more robust, healthier constitution and often accompanied by a late menopause3 and a remarkable fertility (15 children per woman).
Wouldn’t the conscious control of procreation be the continuation of what seems already nascent in our cousins, the complete subordination of reproduction to the metapsychic instinctive program ? While lutein levels could not be measured (to ascertain the absence of ovulation), the typial mood swings during that “day of the month” were not observed, hardly any of the physical signs associated with menstruations and ovulations (in one word, the oestrus !) were observed.
More shockingly, free sexuality without any care for birth control proved the existence in the raw frame of reference, of a fool-proof, natural anticonception mechanism. In Mont-Ramé one subject under close scrutiny (Mr Burger himself) had daily intercourse for a whole year with a certain “Daisy”, with no pregnancy resulting. Then a single intercourse with a very base, low-quality unsophisticated man impregnated her, with a paternity test confirming. The fertility of the male being out of questioning (his 7 children confirming), one would have to assume her cycles had been interrupted for a whole year. But the chances to ovulate in a 6-days window around the window in that precise moment were slim to none. The girl had no desire for baby-making, so we shall consider than the innate contraceptive control came for his previous male lover. That must mean prehistorical men could have sex with anyone without risking admixture.
While ultimately hormones are all commanded by the brain, a number of nerve endings lie in direct contact with follicles in the ovaries, serving as a credible candidate to serve in a conscious or unconscious triggering of ovulation. Similarly, the emission of spermazoa by the testicles during ejaculation is under control of the pudendal nerve, we have known for a long time that orgasm and ejaculation can dissociate completely: none of this is biologically surprising. As the recurrent overpopulation crises in history including in ancient times prove, such mode of regulation was absolutely necessary and the natural conclusion of an ongoing trend along tens of millions of years of evolution.
A mother practicing strictly for about a year will live a revolution of childbearing:
The mother can work as hard as before until the very last days
Children come out with the amniotic sac on their heads, mothers do not lose water. As a result, births happen very quickly, in a matter of minutes
, the bag acting as hydraulic transmission.
The pouch ruptures on its own under the effect of enzymes, and likewise the umbilical cord dries up on its own after a day, giving the usual shape of the navel without the need to make a knot.
Giving birth as never be as quick nor painless: no (wild) animal in creation gives birth any other way. The bellows of agony, various complications, deaths in childbirth, indispositions, mood swings and rejections of the baby are unnatural abberations not even constant among cultures, or in our own. Still today, a fraction of women do no suffer during childbirth… some even orgasm.
It hasn’t aged at all, since medicine hasn’t in the slightest improved its understanding of the viral phenomena, or human biology for that matter. Ergo, everything written below still stands as strong, and time could only add more exemples from molecular and genetic studies.
Pasteur was the first to use the term “virus” to designate the pathogenic action of bacteria he had discovered in the field of the microscope. At the beginning of the century, increasingly sophisticated filters, followed by ultracentrifugation, X-ray diffraction and electrophoresis techniques, made it possible to establish the existence of very small particles, which carry an indefinitely reproducible infectious power, although they lack autonomous vital functions.
More recently, molecular biology and the electron microscope have made it possible to determine and visualize the exact structures of a large number of viruses, as well as the mechanisms of their multiplication and their action on a molecular scale. Thus, the mystery of these infinitesimal beings which had remained hypothetical for so long, associated with so many illnesses and diseases, and even unbearable tragedies such as smallpox or polio in the past, and AIDS today, seems solved.
This knowledge gives us hope to find ways to fight either preventively by acting on the immune system with vaccines, or curatively by directly inhibiting viral activity by molecular means with antivirals. However, the prolonged failure of these techniques in the case of HIV, despite the importance of the technological apparatus implemented, as well as the contradictions that remain between theory and facts, should prompt us to ask ourselves a number of questions.
The basis of the reasoning behind current research is in fact the legacy of an era when the superstitions attached to the fear of contagion and major epidemics were barely overcome. The image that the medicine of that time gave us of the virus, considered a priori as a pathogenic agent, harmful by definition, is not necessarily the only one possible. The current trend is to consider the disease more as an imbalance between the host and the aggressor, giving more importance to the factors likely to decrease the resistance of the organism. A further step would be to look for the meaning of the viral phenomenon in itself, without any emotional connotation.
There are many viruses in the natural world that do not manifest themselves as a nuisance. Even in humans, many viral diseases occur mostly in a frugal or asymptomatic form. In the case of poliomyelitis, for example, serological studies in epidemic settings have shown that nervous system involvement occurs in only a very small percentage of infected persons.
In children, primary infection with herpes virus usually occurs without symptoms. In adults complications are exceptional, with the majority of individuals being healthy carriers. In the different classes of viral hepatitis, there are also a large number of completely latent forms; the benign forms usually end with complete regeneration of the hepatocytes, with restoration of a normal architecture thanks to a remarkable conservation of the reticulum during the course of the disease.
Similarly, the Epstein-Barr virus is detectable in most cases only by hematological and serological examinations, and is found in the majority of African children, whereas it causes Burkitt’s sarcoma in only one case in ten thousand, probably in cooperation with various cofactors; when it triggers mononucleosis, this is usually not serious. The rabies virus itself develops the classic symptoms in some people and not in others, and the reasons for these differences are still unknown.
The situation is similar in animals: avian influenza manifests itself in domestic ducks and quails in the form of coughing, sneezing, and swelling around the beak, leading to significant mortality, whereas it remains mild in other wild or domestic species. The swine influenza virus, considered dangerous, even fatal for young pigs infected by their mothers, is found in pigs from different regions where it is only sporadically accompanied by clinical manifestations.
Many epidemiologists are of the opinion that most viruses are widely distributed in all living species, including humans, but only occasionally manifest themselves by pathological symptoms, under the effect of triggering factors still not well known.
Insofar as the number of invisible forms and healthy carriers turns out to be greater than that of the serious forms, there is nothing to prevent, at least from a theoretical point of view, a reversing of the classical model of reasoning. Rather than considering the viral disease as the natural outcome of the invasion by the virus and being surprised that the virus can be present when nothing is happening, one could postulate that the asymptotic form of the viral invasion is the normal form of a natural phenomenon, the pathological forms being only the result of an accidental evolution due to certain other pathogenic factors.
Moreover, if it is a phenomenon not harmful in itself and belonging to the complex equilibrium mechanisms characterizing biological reality, it should be possible to attribute to it a precise function (a teleological meaning), at the very least a function that is useful to the host, even if this possibility does not yet seem to have been envisaged in the classical conceptions.
To illustrate this in a somewhat trivial way, let us take the case of the rockets used to put satellites into orbit: if the launch fails in one case out of ten, an uninformed observer, struck more by the accidents than by the successes, much less impressive, could think the purpose of the operation is to destroy the satellite and that this purpose is missed in the nine other cases; all the operations conducted by the engineers and technicians would be explained for this observer as well as if he knew their real purpose, apart from the impression of a great quantity of failures and useless efforts; without knowing the real intentions behind the facts he can observe, that is to say, without knowing the satellites can have any usefulness, such an observer would believe it useful to intervene by destroying the satellite with simple explosives rather than to join in the work that represents its launching.
Viruses cause significant problems in about one in a hundred cases. They remain frightening if they have no other meaning than to cause disease. On the other hand, if our postulates were confirmed and if we could attribute a useful function to the viral process on the one hand, while identifying on the other hand the causes of dysfunction causing the accidental danger, a very different direction of research, even therapeutic action, would unveil.
“In the case of AIDS, it seemed a priori that the virus had a harmful effect in 100% of cases. It is significant that the best-placed researchers have come to believe, some ten years after its discovery, that the pathogenic activity of this retrovirus is due more to certain co-factors than to its intrinsic characteristics.
Faced with the general failure of the prophylactic and therapeutic means implemented, and faced with the urgency of the situation, all paths certainly deserve to be explored: the very basis of the reasoning on which medical action is founded, as each time a theory leads to failures or contradictions, must be reconsidered in the light of the knowledge acquired in the meantime and, above all, in the light of the facts that can be brought to light by new experiences.
This is precisely another theoretical model of the viral phenomenon Guy‑Claude Burger, a former mathematician and theoretical physicist, proposes here for the consideration of researchers open to a multidisciplinary reflection. After thirty years of unprecedented experience on genetic maladjustment to traditional food, he hopes to make a modest contribution to the general effort to curb the threatening epidemic and to advance knowledge.
Viruses are generally considered as pathogenic agents, devoid of life of their own and subsisting at the expense of the organisms they infect. The virion (viral particle) attaches itself to the membrane of a cell, introduces its DNA or RNA and hijacks the cell’s genetic machinery in order to reproduce itself.
The new virions spread into the circulating masses and infect other cells. The host’s immune system reacts with varying degrees of success by creating antibodies to stop the process. This is done with a certain delay or failure rate which explains the variable importance of symptoms observed in different subjects.
This process has no other teleological significance than the virus’ multiplication and perpetuation. It is carried out at the expense of a living species that must not, out of necessity, succumb, a fact within limits accounting for a certain balance between the harmfulness of the virus and the resistance of the species.
We know today the sequences of nucleotides of a great number of viruses and retroviruses, as well as the structure of their envelope and the nature of the antigens which allow the immune system to recognize them.
Viral invasion triggers an immune system reaction that results in various symptoms: asthenia, hyperthermia, inflammation of the mucous membranes, catarrh, rashes, etc. In addition, in association with the viral process, especially in diseases of the respiratory tract, the multiplication of pathogenic bacteria is often observed.
Normally, this proliferation is slowed down, for example in the case of coryza, by the bacteriostatic action of the nasal mucus, but this balance seems to be broken by the action of the virus. Similarly, viral pneumonia can lead to bacterial superinfection and various complications, hence the systematic use of antibiotic therapy, although this has no effect on the viral process itself.
In the absence of complications, the viral disease converges spontaneously towards recovery. In some cases, it may leave sequelae (e.g., post-liver cirrhosis) or even lead to death.
The classical means of fighting viral diseases are prophylaxis, vaccination, rest, diet, abstention from alcohol, vitamin therapy, and antibiotics to avoid bacterial complications. More recently, various molecules blocking the mechanisms of viral multiplication, or antivirals (such as AZT), have been used, but with inconclusive results. It can be said in general that there is no satisfactory background treatment for viral diseases.
It is accepted that the evolution of a viral disease depends on the general condition of the patient, but the factors characterizing this condition are not yet clearly established. In a significant number of cases, viral diseases develop in a frugal or asymptomatic form. Since the viral information can remain present in infected organisms for a long time without causing any particular symptoms, the a priori contradictory concept of “healthy carrier” requires definition. This state concerns, for most viruses, the majority of individuals.
Guy‑Claude Burger, a physicist and mathematician, former assistant in theoretical physics at the University of Lausanne, was diagnosed with cancer (lymphoblastic sarcoma of the pharynx) in 1960. For about thirty years, he pursued a dietary experiment consisting of reconstituting a Paleolithic type of diet, in order to demonstrate the influence of a possible genetic maladjustment of the human organism to the data of modern nutrition.
Since the Neolithic period, many artifices have been introduced into food habits, such as cooking, the selection of cereals, the use of animal milk and the manufacture of dairy products, as well as the various processes used in the culinary arts in general. These processes modify the organoleptic qualities of foods so as to increase their palatability (which tends to increase their consumption) and also lead to transformations in the biochemical structures of certain nutrients (oxidation, free radicals attaching themselves to other molecules, heterocycles by heating unsaturated fatty acids, Maillard molecules resulting from the reactions between carbohydrates and proteins, etc.)
However, there is no evidence that the genetic data of assimilation, first constituted in contact with primitive foods, could have been adapted in a few millennia to these new food factors. A possible maladjustment of digestive enzymes, of the intestinal barrier, and of the immune system could explain the appearance of numerous disorders and diseases, as a result of the penetration into the circulating masses of molecules foreign to the functioning of the organism.
Paleopathology corroborates this hypothesis by demonstrating that most of the diseases whose traces we know how to recognize on the bones, did not exist or were very rare before the agricultural and culinary era. These few elements may lead us to wonder about the nature of viral diseases: how would they manifest themselves if organisms were fed according to their genetic programming?
Burger’s experiment consisted precisely in observing, over periods of up to twenty years, a large number of subjects fed according to the Paleolithic model, i.e. exclusively with raw food, organically grown, neither prepared nor mixed, excluding all animal milk and all dairy products, and with a minimum of cereals and selected products. The food intake was regulated by a strict observance of the alliesthetic mechanisms (variations of the olfactory and gustatory sensations according to bodily needs) so as to reproduce as much as possible the primitive conditions of food.
Burger would have noted, under these particular feeding conditions, that most viral diseases systematically present themselves in a frugal or asymptomatic form. The viral invasion and multiplication of virions, however, seem to occur under classical conditions. Indeed, Burger claims to have observed in many cases that infected subjects, even though the disease remained invisible, developed classical symptoms within hours of ingesting traditional foods, i.e. as soon as the foreign molecules of which they could be the vectors had passed into the circulating masses.
If, within the framework of a Paleolithic type of diet, in principle in conformity with the genetic data of the organism, the absence or the reduction of the disorders associated with the viral affections were to be confirmed in a systematic way, the very notion of viral disease would have to be called into question.
A first interpretation would be to say simply that a natural diet confers a better resistance to viral aggression. However, it would also be possible to turn the problem around and stop considering the virus as a pathogenic agent in se, the pathogenicity of the phenomenon being sought rather in certain factors of genetic maladjustment to the unnatural diet.
More fundamentally, one should ask whether the viral phenomenon, which is widespread in the natural world, does not possess a biological function whose teleological significance is still poorly discerned by contemporary medicine, at least when it comes to the human being.
Burger points out in this connection that practically all viral diseases are accompanied by “effluents”: catarrh, perspiration, rashes, diarrhea, heavy urine, seborrhea, particular body odors, etc. Basing himself on these facts of current observation, on the other hand on the current data provided by enzymology, molecular biology, virology and immunology, he proposes the following hypothesis: the viral DNA or RNA would program, in addition to the mechanisms necessary to the multiplication of virions, the synthesis of proteins ensuring the evacuation of certain molecules foreign to the normal metabolism which would have accumulated in the intracellular medium.
It is true that retroviruses have a very restricted genome and that they produce only a small number of different proteins whose functions are already known in most cases. However, it is not excluded that a given protein has two functions, one belonging to the reproduction of the virus, the other to a process useful to the cell, still unknown. Biology has already provided more than one such surprise: many organs have multiple functions, some genes can be read with a shift of one nucleotide and give rise to two different yet functional proteins, etc.
It is no more unreasonable to consider, for example, the hypothesis that a viral protein can, on the one hand, exert a suppressive action on viral replication and, on the other hand, bind to foreign molecules of a given class, in order to ensure their transport outside the cell: viral multiplication would thus be linked to the concentration of foreign molecules, which would explain a self-regulation of the phenomenon such as seems to be apparent from Burger’s clinical observations.
From this point of view, viruses, or at least certain viruses, should be seen as a kind of complement to the traditional immunological system: the latter ensures the synthesis of antibodies responsible for eliminating the antigens present in the circulating masses, whereas viruses would be the agents of a kind of intracellular immunology responsible for maintaining order inside the cells.
In other words, the virus would provide the cell with the necessary genetic complement to recognize and eliminate the molecules that it is not able to control by its own genetics, in particular the molecules that are foreign to the normal mechanisms of assimilation, introduced into the organism by the effect of different environmental factors, in particular as a result of the absorption of food containing molecules that are foreign to the genetic data of metabolism.
The symptoms which appear during the viral process would then reflect the difficulties encountered by the organism to get rid of these foreign molecules, more than a fight against the virus itself.
It is generally admitted that the virus has adapted to the cell by a series of mutations obeying the laws of chance and natural selection. The virion would thus have become capable of attaching itself to certain proteins present on the cell membrane, or even of integrating into this membrane by using, for example, the mechanisms of phagocytosis to surreptitiously penetrate the plasma, and then of hijacking the cell’s genetic machinery to its advantage.
It is also possible to reverse the reasoning and postulate that the cell has evolved genetically in such a way as to proceed to the synthesis of various viral particles, allowing it to transmit a genetic message to the other cells of the organism and to other individuals of the species.
The selection pressure is probably stronger in this second hypothesis (which would thus appear more probable), if we admit, as Burger does, that the information transmitted by the virus allows the cell to eliminate harmful molecules: in a living species whose representatives are in competition, the individuals best equipped in terms of intracellular immunology obviously have more chances of reproducing than the others.
In the first hypothesis, the virus would have acquired during its evolution the ability to bind to certain proteins present on the cell membrane.
In the second hypothesis, the cell would have evolved to endow the virion with proteins capable of adhering to certain membrane proteins, which it would have taken advantage of to ensure this new function.
The ability of a cell to synthesize a protein capable of binding to a receptor, even a distant one, appears for example in the case of hormones or antibodies, and there is nothing to prevent us from presupposing an analogous phenomenon in the case of the virus.
From the point of view of the quantity of information, it seems more likely that a cell can match a new protein to an element whose synthesis it has already mastered, than the reverse, i.e. that a virus can “succeed” by chance alone in synthesizing binding proteins corresponding to proteins which would be in no way related to it.
Similarly, a series of mutations hardly explains the ability of the virion to integrate its own membrane with that of the cell, which requires rather complex molecular mechanisms. Indeed, no process of natural selection can begin before the virus is able to enter a cell to multiply, and no process of multiplication is possible if the virus is not already able to enter a cell. It is difficult to estimate the probability of such an arrangement occurring, but it is certainly very low.
However, this integration phenomenon is immediately explained if we admit that the membrane of the first virus is derived from a cell membrane. This is also perfectly consistent with some virions leaving the cell where their multiplication took place “borrow” their membrane from that of their host, or rather: that the multiplying cell uses its own membrane to “package” the genetic message that it sends to its fellow cells.
The remarkable identity between an important portion of the viral nucleotide sequence and that of the cellular DNA, as it is observed in retroviruses, seems difficult to attribute to chance. On the other hand, it is immediately explained if one admits that the retrovirus, in a more or less distant past, originated from the cell.
In DNA viruses, even if we are not dealing with identical sequences, there is nevertheless a relationship that allows the virus to hijack cellular genetics to its advantage. This “homology” can be explained either by a genetic adaptation of the virus to the cell, or by an adaptation of the cell to an existing virus, or by assuming that the viral DNA is derived, at least in part, from cellular DNA.
Just as the organism knows how to control the multiplication of useful bacteria, for example in the intestinal flora, it is conceivable that it could have “learned” to control certain existing viruses in order to take advantage of them: just as intestinal bacteria are useful to it by their enzymes which complete the range of enzymes provided for in the genetic make-up, viruses were able to provide an assortment of proteins useful for maintaining the integrity of the intracellular environment.
The discovery of an enzyme capable of transcribing retrovirus RNA into DNA defied all predictions of biologists at the time. This viral type suddenly proved to be able to “predict” its copy by synthesizing itself the enzyme necessary to the transcription of its genetic information in the language proper to cellular genetics. Moreover, this fact seemed to contradict everything we knew about the irreversibility of the transcription of DNA into RNA in all living beings.
Such a phenomenon can be better explained if we postulate that the cell, by virtue of a perhaps very old mechanism inscribed in its genetic heritage, has endowed the RNA of the retrovirus with the information necessary for the synthesis of an enzyme capable of reverse transcription into DNA. This allows, on the one hand, the transmitting cell to export information by passing through the classic way of the RNA-polymerase, and on the other hand, the receiving cell to integrate the transmitted information at the level of its own DNA. This reasoning is only meaningful in terms of evolution if one postulates that the transmitted information is useful to the individual and to the species, in accordance with Burger’s hypothesis.
It seems that viroids, short chains of RNA consisting of only a few hundred nucleotides, studied so far in plants, reproduce thanks to the action of enzymes already present in the host cell.
This fact is difficult to explain if one admits that the viroid is of external origin to the cell: it implies that the viroid is able to divert enzymes in charge of other functions in the cell for its own multiplication. On the other hand, they fit perfectly into the logic of an action programmed by the cell, useful to the individual and the species according to the Burger hypothesis.
In this respect, it should be noted that viroids only cause symptoms in certain “sensitive” plants of a species, whereas they are also present in others without causing any harmful effects: the problem of the “healthy carrier”, which concerns the majority of individuals infected by classical viruses, is already present in these simplified viruses. Some researchers consider viroids to be “abnormal” regulatory molecules: since their action is not systematically harmful, there is reason to look for other factors responsible for triggering a pathology.
Nothing prevents us from thinking that these rudimentary viral particles are the result of archaic mechanisms of transmission of genetic information, the secrets of which biology has yet to reveal.
It is generally considered that the virus “hijacks the cell’s genetic machinery for its own benefit” in order to reproduce its own genetic information a certain number of times. This statement is based on the fact that the virus induces in some cases a complete blockage of the cellular machinery, the only genes expressed being then the viral genes.
If one accepts that the expression of viral genes is useful to the organism and the species, one should rather say that certain cells “concentrate their activity on the multiplication of viral information, in order to retransmit it to the other cells of the body”.
The blocking of the normal activities of certain cells does not pose any particular problem for the organism if their number remains limited. Experience shows that such a limitation is indeed assured in the vast majority of cases.
Some viruses, such as the poliomyelitis virus, are known to cause the destruction of infected cells. As in the previous paragraph, it should be noted that the lysis of a certain number of cells dispersed in the organism does not represent an irreversible lesion if their percentage remains below a certain threshold.
The problem is rather to know which factors can cause this threshold to be exceeded: for example, a deficiency in the immune system, or, in accordance with Burger’s hypothesis, an exaggerated concentration of foreign molecules stimulating the multiplication of the virus responsible for their elimination.
If the viral information is supposed to be useful, it does not appear unfavorable in itself that the organism “sacrifices” a limited number of cells in order to ensure their multiplication, as long as the phenomenon remains reversible, i.e. the dead cells can be replaced by operational cells. Still in the example of poliomyelitis, the number of patients presenting irreversible lesions of the neurons (alteration of the nucleus of the cells and irreversible paralysis) is about 0.25%, which is obviously insufficient to be able to consider these lesions as a consequence directly linked to the action of the virus.
It should also be noted that the incubation phase, during which the virus multiplies, is generally silent. In Burger’s hypothesis, the symptoms which appear during the state period should be divided into two classes: those which result from possibly irreversible cellular destruction, and those which are caused by foreign molecules released by the cells into the circulating masses. y
In addition, the destruction of certain cells, as in the case of infections by the herpes virus, could be part of a general programming of the phenomenon including, for example, the formation of papules useful for the elimination of substances rejected by the cells.
The genetic variability observed in many viruses can be accounted for by the diversity of the classes of foreign molecules whose elimination they are responsible for programming. There would be a certain analogy with the multiplicity of the different antibodies that lymphocytes know how to elaborate to recognize the different classes of antigens likely to penetrate the circulating masses. Similarly, the variability of viruses would allow intracellular immunology to cope with the various classes of foreign molecules capable of accumulating inside the cells. It is therefore questionable whether the mutations we observe are not induced by cellular genetics.
The existence of viruses that are obviously harmful to the individual in the plant world can be explained by a kind of homeostasis at the level of the species: the survival of the species is indeed endangered if the biotope becomes unbalanced due to overpopulation. The usefulness for the species seems here to go against the usefulness for the individual. This is undoubtedly due to the fact that the survival of the individual, in the plant world, is much less important for the maintenance of the species than in the animal world, especially in the higher animals where the litters are few.
In this regard, it can be noted that overpopulation causes deficiencies in the humus, which in turn leads to nutritional disorders in plants. Thus, there is already a relationship between nutritional disorders and virosis in the plant kingdom. It is therefore not absurd to think that this same phenomenon could have taken, through the evolution undergone by the animal kingdom, a more elaborate form whose strategy consists in preserving the individual to favor the survival of the species.
The production of interferon during the multiplication of the virion in the first cells, avoiding further multiplication in the other cells, is meaningful if we admit that transmission of the viral information to all the cells of the individual responds to a process of genetic complementation “foreseen” by the organism.
On the other hand, it is difficult to explain in terms of a defense mechanism as the classical model would have it: if such a defense mechanism is possible at the time of viral invasion, it is not clear why interferon would not be synthesized early enough (at a time when the organism is not yet weakened and would therefore be in a better position to defend itself), as is the case for many immunological mechanisms. Such slowness seems to contradict the laws of evolution, whereas the hypothesis of a collaboration between the virus and the cell, useful for the species, justifies perfectly the presence of a regulation mechanism allowing the virus to multiply within the adequate limits so as to avoid that all the cells of the body are infected.
Even if one accepts that another cause of impairment causes the delay in interferon production, it is still troubling that this production can be completed correctly when the two causes are superimposed (viral infection and external cause), in a manner precise enough to keep the number of virions limited to one or a few copies per cell. On the other hand, the hypothesis of a “collaboration” between the cell and the virus fully justifies the presence of such a mechanism, which then appears as a regulatory system rather than a defense system.
The display of certain proteins by cells, which occurs under the influence of interferon (e.g. the p69 protein displayed by pancreatic cells), could have the function of triggering auto-immune mechanisms designed to eliminate cells invaded by excessive quantities of foreign molecules. Thus, letting the virus program the return to integrity of the least affected cells, the immune system would take charge of eliminating the cells that are too severely encumbered and should be replaced. This hypothesis would be confirmed if it could be demonstrated that the display in question is proportional to the concentration of foreign molecules in the cell.
The construction of perfectly structured virions and their expulsion through the cell membrane is the result of a coordinated action, which is very complex if we consider the mechanisms involved. This action is programmed by the viral genome in a surprisingly targeted manner. It seems less risky to attribute its origin to the cellular genome, which has the necessary mass of information, than to a mutation-selection process at the level of the viral particle. This process can only start when the virus is already able to reproduce itself. Insofar as this reproduction can only take place in the cell, it is difficult to see how the phenomenon could have been initiated.
The fact that the viral information is stored in the cell and hidden, with the possibility of being reactivated, seems more logical if one postulates that it is useful information, allowing the cell to ensure the evacuation of certain harmful molecules, even if it means reactivating the process at the moment when their accumulation becomes detrimental.
The classical view of the virus as a simple pathogen would suggest that virions and their genetic content are totally destroyed after recovery, at least in the most resistant individuals. However, the persistence of viral information actually is the rule.
If it has not yet been possible to identify the factors likely to trigger the reactivation of the viral process, it is perhaps precisely because they involve not only the biological data of the virus and the cell, but also the biochemical properties of molecules whose existence has not been taken into consideration until now.
The bacterial infections that we often see associated with viral diseases could be explained not only by a weakening of the immune system, but by the presence in the circulating masses of foreign molecules rejected by the cells.
Two hypotheses are therefore possible: either these foreign molecules weaken the organism and open the way to bacterial invasion. Or the multiplication of certain bacteria would also be programmed by the viral information in interaction with the genetics of the organism.
This second explanation is not absurd: the foreign molecules whose presence we postulate escape by definition from the mechanisms of assimilation as well as from the vigilance of the immune system, since they have been able to reach the interior of the cells without ambiguity; their elimination thus requires mechanisms which do not belong to the organism itself, for example bacterial enzymes able to degrade the undesirable molecules.
This model of reasoning is in agreement with what we know about the bacterial flora: there too, the organism seems to have been able to “domesticate” bacteria whose enzymes allow it to degrade molecules that escape its own enzymes, for example carbohydrate chains like cellulose.
Thus, the virus would induce not only the processes necessary to maintain intracellular integrity, but also the multiplication of bacteria capable of degrading the waste products rejected by the cells. The apparent pathogenicity of these bacteria could be attributed less to the virulence of particular strains than to an excessive level of target molecules in the circulating masses.
Apoptosis, the process of natural cell death observed, for example, in T4 lymphocytes in the presence of HIV, would have the following meaning in this conception: the virus would program the suppression of lymphocytes in charge of recognizing classes of bacteria, whose enzymes are required to degrade the molecules rejected by the cells, so as to favor the multiplication of these bacteria. The viral genetic information would thus program the transport of foreign molecules out of the cells while bacterias multiply likely to rid the circulating masses of them on the other.
Under the effect of an exaggerated concentration of target molecules, especially when molecules of the same type are brought in daily by unsuitable food, it would make sense in this hypothesis that apoptosis exceeds the correct limits, and the immune system enters a state of apparent deficiency, letting all sorts of pathogenic elements develop freely.
Autoimmune mechanisms, triggered by the presence of food-borne antigens also bind to lymphocyte membranes, could complicate the phenomenon and aggravate the destruction of these cells.
The more or less severe evolution of the viral process in different individuals can be explained by a more or less important accumulation of foreign molecules, according to the different food anamnesis.
The symptomatic form that it takes more regularly in the human species would be due to the fact that the food proper to the civilization has considerably moved away from the primitive food which could have determined the evolution of our genetics, and it is unlikely that in a few thousand years the human organism could have adapted genetically to all the new molecules brought by the agricultural and culinary artifices established since the Neolithic.
A virus as dangerous as SIV hardly causes any symptoms in monkeys living in their natural environment, nor even HIV in captive chimpanzees fed in a natural way. Since the regulation of viral multiplication depends on the presence of foreign molecules in the body, it is to be expected that additional intake of the same molecules through conventional foods would disrupt the process.
According to Burger, the consumption of certain foods by infected subjects during the incubation period would cause an aggravation of the subsequent symptoms, for example in viral hepatitis. From this point of view, it is understandable that the diet prescribed regularly by family doctors to patients with influenza, coryza, hepatitis, etc., has had enough effect to be maintained in the medical tradition.
The popular wisdom which attributed a utility to the diseases of children, for the majority of viral etiology, is paradoxically justified: the organism equipped with the complements of genetic programs brought by the various viruses is better armed against the harmful molecules likely to invade its cells during their existence. This calls into question the fundamental meaning of vaccinations: their usefulness would be to avoid viral invasions that could have serious consequences in the classical food conditions.
On the other hand, if Burger’s hypothesis were to be confirmed, there would be reason to fear that the absence of the genetic complements provided by common viruses would deprive individuals of the mechanisms provided to ensure the maintenance of the integrity of the cellular environment, with the risk of aggravating degenerative processes and compromising various functions of vital importance.
The expansion of certain viral diseases in wild animals can be explained by changes in the environment, the cultivation of cereals or other mutated plants introducing appreciable quantities of new molecules into the natural food environment (proteins produced following mutations in wheat, for example, accumulating in the body of rodents, then in that of the fox, causing the activation of the rabies virus, which was already present before without causing any particular problems). To this could be added the influence of molecules introduced by industrial waste and pollution.
The relative failure of so-called antiviral molecules can be explained by the difficulty of counteracting vital processes programmed by genetic means. The interactions between the viral genome and the cellular genome take place in the cell nucleus and respond to precise mechanisms, so that it is very difficult to inhibit them without harming the cell at the same time. Such processes probably include self-regulatory or substitution mechanisms intended to guarantee their action, the rebellious nature of which may seem paradoxical as long as their biological purpose cannot be defined.
The case of oncogenic viruses occupies a special place; they may always be harmful. However, the multiplication of cells can be useful for various purposes, if only to compensate for cellular destruction due to some cause. It would therefore also be possible to envisage that these viruses provide useful information to the organism, even if this means that they can lead to disastrous results under the effect of certain cofactors.
The Epstein-Barr virus only manifests itself as a sarcoma in a very small proportion of infected children, and only in Africa. In addition to genetic predisposition, it would be appropriate to investigate, on the basis of Burger’s hypothesis, the presence of certain foreign molecules reaching a particularly high concentration, resulting for example from the dietary habits of young Africans.
As far as HIV is concerned, it has been admitted that almost all infected persons should develop severe symptoms. Indeed, the evidence so far has confirmed that, with very few exceptions, HIV-positive status will result in a fatal outcome. These facts seem to contradict the previous points. However, the same or similar retroviruses have been found in recent years in many wild animals, which do not seem to show any particular symptoms.
The best researchers have come to believe that the pathogenicity of this virus is due rather to some as yet unknown “co-factors” than to the nature of the virus itself.
In Burger’s hypothesis, these co-factors could be the molecules that the virus would be responsible for programming the elimination of, present in much greater numbers in human organisms than in wild animals: the latter feed mainly on natural foodstuffs to which their genetics have been able to adapt since time immemorial, whereas humans regularly absorb traditional foodstuffs that did not exist in the primitive environment, and to which human genetics have hardly had time to readjust.
It is therefore to be feared that certain foreign molecules will have the opportunity to accumulate in human cells at concentrations that have never been reached in the history of the species. The viral processes in charge of programming their elimination, in a primitively silent way, would thus be confronted with an unforeseen situation: the abundance of target molecules would disorganize regulatory mechanisms that ensure their proper functioning and would lead to the appearance of dangerous “opportunistic infections” as a result of an exaggerated multiplication of the associated bacteria.
It remains then to explain why this retrovirus, which was perhaps part of the genetic heritage of humanity without signaling itself, as it is the case in animals, would be suddenly out of the shadow of the cellular nuclei to cause a serious epidemic. Among the reasons to be considered are the changes in eating habits, which have been considerable in the last decades, especially in the Third World countries, where Western eating habits spread quite suddenly, as well as new causes of contagion. Once viral multiplication has been triggered, the virus could only improve its “performance”: the most contagious virions and those causing the most mucous lesions are the ones that multiply preferentially.
In addition, organisms no longer in possession of the virus or in which it was more deeply inactivated, had the time to accumulate a particularly high quantity of target molecules. This would explain the particular violence of the viral process, further increased by the daily intake of foodstuffs carrying molecules of the same classes.
A new theoretical model, in a field as complex and charged with emotional factors as disease and contagion, can only be verified with sufficient hindsight, through the coherence of the reasoning to which it provides the starting point, and above all through the facts.
Unfortunately, it is not easy to obtain the publication of new ideas not yet endorsed by the scientific corpus, even if it is only a matter of submitting them to the criticism of specialists. Burger therefore invites all interested researchers to criticize his proposed model in the light of their theoretical knowledge, and all practitioners to observe whether the presumed relationships between patients’ diets and the evolution of classical viral diseases correspond to possible predictions. He would be glad if those who observe either significant contradictions or concordances could take the trouble to communicate them to him.
If Burger’s viral model proves successful, it could open up a new avenue of research, especially in the field of AIDS. It would not only be a matter of looking for a vaccine or developing antiviral molecules to deal with the most urgent problems, but also of identifying the molecules of food origin potentially responsible for disrupting the viral process.
Dietary measures applied as a preventive measure could consequently improve the future prospects of current seropositive people. The daily intake of foreign molecules may also play a role in the regulation of the viral process. A correction of the food hygiene could, in this hypothesis, improve the fate of the persons already contaminated, and perhaps limit the evolution of the symptoms even after their appearance.
It is regrettable that no epidemiological research has been done so far to establish the possible existence of a relationship between the dietary history of HIV-positive persons or the daily diet of AIDS patients and the severity of symptoms.
Furthermore, the identification of food-borne xenobiotics could lead to a better understanding of the cause of many dysfunctions affecting metabolism or other functions involving biochemical mechanisms, such as the transmission of nerve impulses, DNA replication, etc. Certain proteins contained in wheat gluten (gliadins) seem to aggravate the symptoms of schizophrenia, various Maillard molecules have been shown to be mutagenic, and there are certainly many pathogenic factors still to be discovered in this field.
In the same perspective, the heuristic proposed by Burger would lead to a more systematic search for food-borne antigens involved in the genesis of autoimmune diseases. The recent discovery of a peptide in cow’s milk, apparently responsible for the reversal of the immune system against the B cells of the pancreas, and opening the way to juvenile diabetes, as well as various experiments on rheumatoid arthritis, improved in some 80% of cases by a diet excluding cow’s milk, wheat and their derivatives, go in the same direction.
To replace the prevailing germ theory, from the basis of our experiences and the observation of animals and the circumstances in which diseases arise, Mr. Burger created a new theory of diseases, pathogens of all kinds (viruses, fungi, bacteria and parasites) and their relation to health. Each layer of complexity in the realm of life, is under the genetic and molecular command of the layer above. We do not fight “pathogens”, we do not cohabit with them in symbiotic relationships like equals in a partnership.
Our genetics commands them, using them to improve the molecular order of our organisms: cleaning us, repairing us and sharing genetic information in our stead. We claim that microbes and multicellular parasites alike, serve the need of “higher” species, both animal and vegetal, keeping them fit and genetically strong and partaking in the horizontal transfer of genetic material throughout the whole ecosystem, as recognized today.
Parasites are supremely varied and numerous, because they fill a crucial role in ecological equilibriums. They are a powerful agent of natural selection on a molecular or physiological level. However, if the role of parasites and viruses is to weed out the weak and diseased, the degenerates. Keeping the population fit. The question is, how many “weak” individuals is a natural population—in a pristine natural setting—supposed to carry, that would determine its mean parasitic load ? At what price for any individual is this balance achieved ? In our experience parasites (such as tenias) are always regulated. A few days of disciplined instincto suffices to expel in mass in feces, in the form of crawling neatly packed balls of living (or dead) worms.
Pets and cattle show similar (though less potent, in the sense that our levels of cooking kills them) mechanisms of molecular exchange when provided with equivalent natural molecules, gradually eliminating culinary toxins from their body. The same story of rapid and complete recovery ensues: invariably an adequate wide food range eradicates parasites and symptoms.
Epidemics indeed can be seen in the animal world, and animals do fall prey to parasites, to various levels, while others don’t… but still carry the pathogens. The scientific world doesn’t understand those variations of sensibility and blame genetics whenever it is convenient, to force reality into their prey-predator model generalized to all microbes, parasites, viruses, etc, like forcing a square peg into round holes.
Maybe we ought to consider broadly two kinds of species, the “evolved” ones whose individuals treat parasites and viruses like mere tools, and primitive ones which rather discard the unfit individuals themselves instead of the waste they carry, because of their very limited capacity for homeostasis. Homeostasis has a complex relationship with complexity, metabolism, intelligence and lifespan, but overall complex creatures capable of learning typically belong to the first category, while simple, instinctive organisms rather to the first. A more direct criteria might be the state of advancement of adaptive immune systems.
“Primitive” species typically breed very fast, in large numbers, and typically live short lives. Bacterias being a good exemple, most plants too. Individuals do not matter one bit, because a handful of them can recreate the whole popuplation in a year. Ecologically, animals with higher lifespans are more likely to show advanced immune systems as losing individuals for a species (always of slow-breeders) is dramatic, an important blow. Maintaining life and body integrity during a long but very active life despite the constant breaking down of cells and proteins, requires either/or an efficient physiology limiting the turn-over rate to a minimum or incredibly fine-tuned cellular biology, completely reverting the accumulating damages (internal and external).
Often, we would need to assert on a case-by-case basis. But at the very least, we are positive that the appalling frequency of epidemics and parasitosis in higher vertebrates like foxes, boars, rabbits, or at sea Greenland or Pacific sleeper sharks or dolphins, can only be explained by a general pollution in the environment, a priori on the whole planet.
Simulacra (as defined by Konrad Lorentz, displays whose real purpose is different from the real one) go completely out of control. In the case of chimpanzees simple brawls aimed at deciding new hierarchies, can change into seemingly organized murders of the kind only humans are capable of.
All animals, bar humans, should by definition all practice our diet constantly, given that animals can only ever guide themselves with their senses alone and do not in theory have big enough of an intellect, to contradict their instincts. Regardless of specifics, what they eat in their natural environment is the natural diet by definition, and the only one life has even known since humans, and more importantly the extensive pollution we brought to this world.
Consequently, we should be in the position to explain how behaviors and morality an animal should be expecting to have, what makes sense and what doesn’t, and what could explain observed abberations. No one else, has the conceptual framework for this, because no one else has even a beginning of understanding health as well as the nature of the mind. Wihtout that understanding, one simply cannot argue from a completely rational ground, what ought to be natural or pathological, be it in men or animals.
What we have been observing for decades is the alteration of behaviors with any form of processed food (the more processed the worse) compared to raw paleolithic food, so, imitating wild animals. For us humans, it meant imitating chimpanzees. The entire world is afflicted afflicted with low levels of pollution, plastic bits or otherwise, from the bottom of the ocean to the depth of the Amazonian forest, although the later case is yet to find a proper explanation. Molecular stress (any unnatural amount of molecules not found in the species’ natural environment or diet) almost universally make animals less alert yet more excited, increasing levels of breeding behaviors and exacerbated secondary sexual character traits like territorial dominance; male aggressivity and female passivity to non-functional levels.
Until the complete natural environment (in terms of food range at least) of a population has been reconstituted to a significant degree within a perimeter, speculating about ape nature is moot. The likes of Wrangham find an obvious perverse pleasure in tarnishing the public image of animals, almost on a sexual level. The mainstream public is simply too eager to accept any pseudoscientific theory relieving them of the guilt borne of comparing harmonious functional primate societies with our highly dysfunctional, mentally insane societies. It is easier to blame genetics and nature, than to put the effort into correcting one’s lifestyle and culture.
Because natural selection is mostly (not only) concerned with the immediate survival of a lineage (not even a species), it might produce all sorts of disparate combinations of personality traits, instincts and cognitive abilities. However in the grand scheme of things, if intelligence could develop in ways barring access to higher forms of love and empathy, or spiritual awareness, such paths are evolutionary dead-ends, as they directly contravene the conditions for higher sociality, which does imply empathy.
Closer to home, the exemple of cats “playing with food is a bad one, as we know it is not natural to them either. Cats only display such “cruelty” because they have the instinct to hunt, but their processed diet renders them incapable to eat live prey. Predators either eat or leave preys alone, not avoiding needless expenditures of energy. Lions may catch a prey and delay the killing but only to let his offsrping learn the trade without risk: while a unpleasant fate for the animal, there is no cruelty in this. This demonstrates amorality as opposed to imorality.
Objectively insane or mean behaviors (such as interspecies rapes, or rapes in otherwise empathetic, social animals) are extremely rare in nature, but they have been reported, in some species more than others. We believe they nearly always contradict basic evolutionary principles, and we intend to prove they all stem from chemicall-induced insanity.
Bottom line, we cannot take for granted anything we see anymore in the wild, from behaviors to epidemics to ecological dynamics. We created a fallen world.
Nature, for a century, still existed untouched in a few places, and the boundary between the sullied and unsullied was a lot easier to tell. It was exponentially easier yet 60 years ago, to observe in a natural state, in terms of behaviors, diet and health.
Today thanks to industrialization the encroaching of men on nature has become nearly total. Pristine nature has become nearly extinct, human depredation (through logging, pollution and poaching) has become extensive to the point that we find significant concentration of plastic bag in the Arctic circle, thousands of miles from any inhabited land, and at the very bottom of the world. Today there is no place however remote, which could by default place observations out of reach of the taint of human interference. Until proven otherwise and explained by a theory based on evolutionary reasoning and our experience of natural food as observed on men (and a few animals as well), no “fact” is totally beyond our suspicion. Facts lie.
While from the 1960s to 1980s a deep ecological (close to Rousseau-ist) conscience among scientists arose, it seems from 90s the tendency among many scientists and the public opinion reversed, with disastrous consequences in the long run. A slew of so-called experts of the likes of Richard Wrangham) endeavored to undermine those much warranted efforts. Their collective strategy (no conspiracy theory, this required only a common fundamental hate for nature and dishonesty) consists in two principles:
Falsely representing as “natural” newly data found in thoroughly damaged environments.
Providing a hugely biased interpretation and abunding in extremely argumentative theories whose only purpose is to warp public perception of nature into a disease-ridden hellscape of violence and rape.
For a few decades science has had a remarkably progressist role on the ecological front as a public institution, furthering the idea that men’ issues wereAlthough on a lesser number of patients, we observed the same rapid and complete recovery of animals, mainly farm animals and pets. Invariably an adequate wide food range ensured the swift eradication of parasitoses that no other methods (sometimes even resisting chemical treatments) could get rid of. Typically in the form of crawling neatly packed balls of living (or dead) worms, in feces, showing up in feces, most often no more than a mere few days after stopping cooking completely. the product of history, civilization, not of its nature, and that primitive cultures were mostly devoid of our unrest, violence and many of our diseases. We can see a gradual reversal of those considerations from the 90ies, now depicting apes not as wise ancestors living in a constant paradise (which they are, or should be) but as poor creatures bent constantly striving to survive in a dangerous place among predators, pathogens and most of all… their own peers. Human violence and sadism is now increasingly explained as a genetic legacy from violent ancestors.
Mountain gorillas have remarkably low levels of either tooth decay or diseases, despite a rate of inbreeding superior to anything we know in human population, to the point of all mountain gorillas being as closely related as half-siblings (35% homozygosity). Yet it has been proved that it didn’t seem to impact their health. But for our instinctive standards they still are very sick, because of a litany of symptoms that demand explanations, and are typical of cooking-eating people with a terrible diet. Overall, from the perspective of a modern conscience those mountain gorillas fare surprisingly well. Yet this amount of tooth loss and over-inflammation (umbrella term for a whole bunch of symptoms) to the point of leaving abscess and attacking bones cannot make sense without a sustained metabolic stress caused by a chronic state of dietary imbalance.
Gorillas eat small leaves, stems, fruVirungaits, bark, and buds, and occasionally small invertebrates like ants, worms, termites, and larvae. The Western Lowland Gorilla subspecies (Gorilla gorilla gorilla) eat the fruits of more than 100 species of trees, 97 of which produce fruits seasonally. About 67 percent of their diet are fruits, 17 percent are leaves, and the rest are seeds, stems, caterpillars, ants, and termites. Sometimes, they enjoy aquatic plants. The other three gorilla subspecies consume much fewer fruit. The mountain gorilla (Gorilla beringei beringei) lives in areas of high altitude above sea level, so their habitat lacks fruit trees. Therefore, they consume leaves, stems, and buds of 142 species of plants, representing up to 86 percent of all their diet. Only 2% are fruits (three kinds of fruits), 3% flowers, and 7% roots and invertebrates.
Virunga mountain gorillas (those eating less than 2% fruits) have neared extinction several times in the last three centuries (Cross River mountains are nearly extinct for this reason). Along with their numbers, their living space dwindled too and fragmented, especially due to the years of civil unrest in Congo and in the late 1960s the cultivation of Pyrethrum halving the Virunga Conservation Area, slashing their population further and confining them to isolated deserts.
Simply put, to get the same amount of calories, Virunga gorillas eat several times more than Western Lowland gorillas which get as much fruits as they want, or close to.
An analysis of chromosomal sequence sharing within individuals (genomic tracts of homozygosity) provided insight into recent ancestry and a measure of parental relatedness. Within mountain and eastern lowland gorilla individuals, chromosomes are typically homozygous over one-third of their length (on average 34.5% and 38.4%, respectively) (Fig. 2B), much higher than in western lowland individuals (13.8%) and exceeding even the most inbred human populations (18). We observed longer tracts in the eastern species, particularly in mountain gorillas, and a clear distinction in tract length distribution between eastern lowland and mountain gorillas (fig. S15) (15). Very long tracts (2.5 to 10 Mb) are particularly indicative of recent inbreeding, and homozygosity on this scale in mountain gorillas exceeds not only that in other gorilla populations but also that observed in the Altai Neandertal (19), consistent with parental relatedness equivalent to that between two half-siblings (19). These data suggest that mountain gorillas may have experienced several recent generations of close inbreeding.
As judged from the date when inferred Ne began to differ, the divergence of eastern and western gorillas began at least 150,000 years ago, but a more direct analysis using male X-chromosomal sequences suggests that they exchanged genetic material until around 20,000 years ago (Fig. 3B) (15). Given that this also coincides with a notable decline in western lowland gorilla Ne (Fig. 3A), it may be that environmental changes during the Last Glacial Maximum (26,000 to 19,000 years ago), when dry savannah replaced tropical forest over much of the Congo basin (21), triggered a collapse in the western population and complete separation of the two species.
Researching for calories intake would be tedious (and too variable) so I’ll keep with food weight instead: in our experience men normally eat (at most) between 1.7 and 2kg per day. Let us take a weight of 55kg (mine), making up between 1.8-3% of my body weight.
Mountain gorillas (males) on a 15%-fruits diet eat on average 18.8kg4 (I wonder how they managed to assess that though !) and weight on average 163 kg (those figures come from the wild since they do not survive in zoos.). Which means a rather robust estimate of 11% of body weight (up to 17% considering an upper limit of 190kg), proportionally three times our quantity. Which makes sense considering they have no more than five fruits in their whole environment, and never at the same time, while it’s about the amount we have in the fridge in any given week.
Western gorillas in nature weigh substantially less (146 against 157 kg in captivity. Males in captivity however can reaching extremes between 275 kg and 310 kg. We find statements claiming mountain gorillas in the wild eat more than their frugivorous counterparts yet it doesn’t appear to hold true for captive western gorillas, which are reported to eat the same amount per day: 18 kg ! 12% of their weight per day is absolutely unjustifiable given a mostly frugivorous diet.
In all likelihood… zoo (Western) gorillas eat two or three times more than they should. As a general rule: the naturality of diets can never be assumed beforehand, only verified. Reserves and national parks are slightly too heavily “managed” and the impact this fact has on animals’ health depends on whether or not they feed them, information typically not mentioned. In some cases we do know reserve gorillas are fed, and the same toxic poisons of their zoo counterparts5.
It seems that zoos still give them a lot of vegetables so counting calories is not possible, but their wide variation in body weight along with the horrible preparations primates are given in zoos does shine some light as to the potential cause of diseases. They are eating cooked food, still today.
In general, zoo primate diets include commercial monkey biscuits, high-fiber, old world and new world primate pellets, or canned primate diet. The appalling poverty of their diet and the fact they eat cooked food automatically discard any study whatsoever done on animals fed by humans. The same can be said of any primate species under human custody. Depending on the place and number of animals concerned, reserve animals may or may not be fed. However they will suffer regardless of a lack of variety, as reserves struggle politically to maintain both resources and acreages.
Forest fragmentation is a dire issue for animals, as it inhibits the natural circulation of seeds and fruit species, sometimes forcing apes to scout in cultivated areas for more food, leading to aberrations. While gorillas are very flexible and their diet reflects the available food, regardless of subspecies the more fruits available the more fruits they will seek preferentially.
Due to both human encroachment into their original habitat and most likely the effect of climate change since the end of the last Ice Age (at the Younger Dryas circa 10ky BCE), those gorillas were forced to migrate into widely unsuitable climatic zones with very little fruit.
Experts argued that the over-eating we observe for mountain gorillas is explained by the low nutritional quality of the plants they have access to. But the fact they are so specific about their food point to a different cause: gorillas are not herbivores, the same way pandas actually love fruits and meat but rely on bamboos out of necessity without their digestive system being adapted to it (remaining the same as that of their cousins, the bears).
Despite the demonstrated great pickiness of apes in matters of food, the idea that their well-being might be negatively impacted by their current possibilities of alimentation has not pierced the unbelievably thick skull of the people in charge of zoos and reserves. Chimpanzees and lowland gorillas on the other hand having as many fruits they want are as versatile, can make-do with just a few fruit species sometimes depending on the season, without sign of lacking. They do not have to constantly collect dozens upon dozens of different innutritious fibrous plants—and be very specific about which parts.
Literature6 show that gorillas do not eat as much fruit as chimpanzees but less out of choice, as because of the environment. While placed in similar environments, they eat similar amounts of fruit. To think diseases are natural because apes should be fine with some fruits is nothing butretarded (not nearly as much as us) is eminently dishonest. We evolved along millions of years along a certain range of environmental variation and this cannot be offset in a mere decades, or centuries, or even tens of thousands of years.
Outside that range we may survive but in suboptimal conditions. Studies seem to agree that mountain Gorillas came to these altitudes due to climate change since the last ice age, an unfortunate event as a duration of 10 000 years pales in comparison of the millions evolution requires regards to the digestive system and dentition.
In those conditions, to conserve animals in their “natural” environment is foolish and even criminal in case of endangered animals (as are most apes). The environment changes faster than animals can adapt ergo if we want pandas and apes to not die, we should relocalise them in rich environments, with a wide range of fruits and prey.
Jane Goodall wrote how chimpanzees would seek out the staff’s feces to eat them. In our experience, animals are very attracted to feces from humans eating cooked food, due to their abnormal scent, and ingesting them excites them considerably. Ditto for any human waste, and cooked food.
Do you know if that still happens ? Would it even be recorded ?
Is it still common (as it once was) for staff members to share their leftovers with apes ?
If animals are fed anything processed, they will suffer the same consequences as men. For each particular case of disease related, one can easily point at glaring disturbances in the environment. The single, overarching cause, being pollution of any kind, plastic heavy metals, industrial compounds or anything men can’t help but “developed” Üntermensch can’t help but diffuse wherever they live. The case of the Mayaro Virus in French Guyana illustrates this sadly well.
“True” nature has become very scarce, it might not even exist anymore in Africa, let alone in the rest of the world., explaining both the prevalence of parasitism and unadaptive behaviors as both indicate a poor constitutive health caused by impoverished habitats.
We must investigate in each case:
What exactly (or on average) is the average food diversity available in most reserves or populations studied ?
What proportion of domesticated vs wild varieties can they access ?
The bonobo has no significant predator, apart from humans. The leopard does not significantly impact the demography of bonobos, which is entirely regulated by food availability. Since our intelligence has reached a certain threshold, say 1000 cm³ and our mobility with it, the pressure of predators (already low) has completely disappeared. On the contrary, the prey was able to become the top predator with disconcerting ease: nothing is too big for thirty spears, a dam or a forest fire. However, nature did not suffer before 30,000 years and wheelbarrows. Diseases… do not exist with instinctonutrition, but that is the theory. What about observations ?
Conscientious, intelligent observation demonstrated that humans are responsible, directly and indirectly for halving the mean lifespan of chimpanzees, from about 32 to 15. It applies to apes at minima, but without doubt to all animals for which predation has become negligible relative to their natural life expectancy7.
In 2001, a meta-study summed up and averaged the data concerning mortality over 5 different sites and representing “represent 3711 chimpanzee years at risk and 278 deaths”, and claimed to have verified statistically an enormous mortality among wild chimpanzees compared to their captive kin (giving a lifespan of respectively 15 and 35). Many of these sites have been the place of poaching and heavy epidemic onsets, both brought by humans.
Though not glossing over the very debatable merit of including deaths seemingly so unnatural, the researchers answered with a dumbfoundingly candid answer, claiming that since primitive humans have cohabited with and hunted apes a few times, the deaths due to epidemics and poaching alike should be considered natural. They considered ebola, as natural.
And as the icing on the cake, I shall remind you that among these sites, Gombe (Gomba in the text) saw the most violent and vicious chimpanzee war ever observed. That alone should have been a red flag, if not for the continuous effort of Wrangham and his ilk to convince the world that bloodlust and rapes had been passed down to humans by chimpanzees. They did acknowledge a significant variation among sites, without detailing. Nor do they detail the different means obtained by removing different sources of mortality. So we were led to conclude that apes led unfairly short and painful lives in the wild, from which they were saved by zoos and closely monitored national parks.
Then in 2017 the truth came out8. It was shown that, besides predation being indeed a non-factor in mortality, under good ecological conditions: The average lifespan reaches similar levels as it does for captive chimpanzees (32 years) and human hunter-gatherers. Considering captive apes’ constant medical care and humans’ higher biological lifespan, that says a lot. Infant mortality (first year) was halved compared to Gombe, Mahale, Taï and Bossou, four of the five sites in the previous sample. Epidemics and poaching were absent.
In conclusion the authors rightfully attributed this massive health differential to the rich environment the Ngogo community profited from, compared to their neighbors at Kanyawara, and the other four sample communities. Better and richer food makes for a stronger immunity, regenerative abilities and assumedly lower stress levels, making abnormalities like the Gombe war unlikely. Apes just should not die from infections, degenerative issues or tumors (the most common causes among captive ones9). For highly evolved long-lived critters with no predator and inhabiting rich environments, the main source of death, beside infant mortality, should always be slower or faster aging due to individual genetic differences.
There is evidence however, that suggests the environment in Ngogo still isn’t perfectly pristine. If wide-sweeping epidemics are unknown, diseases aren’t10. If deaths due to diseases seemed both rare and hard to prove, many individuals, especially approaching a relative old age (36, despite the older chimp reported reaching the 55), have been shown suffering from respiratory ailments common in all zoos. While individual genetics might play a role, it has been proved that non-lethal but chronic ailments act as sources of metabolic stress, reducing resilience to wounds falls and accelerating aging. The same stress that is known to increase inter-individual aggression, sexual violence, infanticides, etc. Said lethal aggression (mostly toward other communities), without reaching Gombe’s levels, seems important in Ngogo.
From these animals’ heightened stress levels we can expect environments of lower integrity to foster all the above causes of deaths: adult diseases, excess infant mortality faster aging, con-specific killings or otherwise antisocial instincts going overboard. And since all of these do certainly exist to some (very) limited degree, telling apart the natural, genetic components for each rate from the artificial influence, is impossible without a proper theory of what a natural environment means on a molecular level.
As the very fact of observation (and the constant monitoring, isolation or care) proves, a pristine nature can never be assumed. Ergo, without knowing beforehand about the natural laws of alimentation, the dependency on observations makes researchers automatically misconstrue the natural state for a given species.
One third of all gorillas and chimpanzees have died from the Ebola crisis since the early 1990s, around 130 000 deaths for each species according to estimations. And Ebola has been heavily linked to deforestation.
The world’s remaining wild apes are being increasingly forced into isolated pockets of forest, which impedes their ability to forage, breed and to hide from hunters. There is also a growing body of evidence linking deforestation and subsequent changes in climate to the spread of Ebola and other infectious diseases… The virus is even more deadly for other great apes than it is for humans, with mortality rates approximately 95% for gorillas and 77% for chimpanzees. Current estimates suggest a third of the world’s gorillas and chimpanzees have died from Ebola since the 1990s.
We are not talking about small, arguably “primitive” animals. These are the peak of vertebrate evolution, bonobos being the closest thing to us. It’s been observed numerous times that an excess of something natural, would often lead to symptoms not too dissimilar, to eating the processed or cooked version of that thing. In particular excess domestic meat, and also fish (including wild ones). The lack of varieties (of a wide enough food range) weakens the body all the same, with upsurges of autoimmunity such as joints hurting.
It is no secret that if an animal will instinctively smell around for hectares in order to find the one thing he prefers, never having that luxury is bound to bring some health issues. We are inextricably tied to the environment we evolved in, on a physiological level. While they can tolerate it, our wider evolutionary line apes did not evolve for dry areas like savanna. But too much aridity will drastically reduce the biodiversity, and available food range, which, as animals lack the foresight and intelligence to move across a whole country to get back in the forest, trap them inside a constant state of weakness… Which parasites and viruses are designed to screen out.
Satellite map of world active fires
Then we have the proximity of anthrax outbreaks, and drought. Fires are visible from space in the infrared with 102,738 fire alerts in the DRC in 2020 alone indicating that Africa is literally burning to man-made fire for agricultural purposes, as you read those words, all year long. How much of that aridity is due to the presence of man ? Even the Sahara only took off to the extent it does today, around the Egyptian predynastic period, 5,500—3,100 BC. Before, a lush climate reigned, hosting lions, crocodiles, hippopotamus and other animals. If this could be said of the desert, then how much more humid were equatorial regions, when it is an established fact that the presence of forests increased rainfall11 ?
Infrared detection of fires in Angola
The high temperature and moisture of tropical rainforests cause dead organic matter in the soil to decompose more quickly than in other climates, thus releasing and losing its nutrients rapidly. When farmers cut down tropical rainforests and use its soil to try to grow crops, they find little success because of the poor nature of the soil.
Because of agriculture and the incapacity of people under cooking to savor original super-food (the nectar and ambrosia of the gods), we destroyed huge swaths of sensitive forest areas where before animals and pre-cooking humans could find all the variety they would need. Besides, forests maintain wetter climates not only locally, but globally as well, helping cloud formation by emitting in the atmosphere molecules which become kernels of nucleation. The same clouds also reflect more light from the sun, reducing temperature.
While carnivores (the top of the food chain) concentrate the pollution from their prey, grazing animals (herbivores) however seem to be more dependent12 on plant variety (no wonder) for their health. The lack of proper food and water understandably induces deficiencies, and causes anthrax, pneumonia, which for instance killed 23% of elephants in Lake Manyara National Park in 1977. and other Hence in places plant diversity can dwindle to almost zero, conditions become wholly inhospitable for life13.
In 2004 has been published Lethal aggression in Pan is better explained by adaptive strategies than human impacts by Wrangham & Wilson, according to which the regularity and pattern of killings in chimpanzees reveal a vicious, “demonic” (the term is theirs) and violent male nature, bent on treating females like resource, and resort to bloodbath whenever it seems advantageous for their selfish interest. Are chimpanzee males driven by the love of their group and the wish to get along like we would expect (a bit naively perhaps ?) any evolved critter (such as intelligent aliens) to behave ? They claim that chimpanzees’ killing rates do not differ from estimated hunter-gatherers’ tribal conflict frequency.
Except the frequency in primitive societies has been very overestimated. Lowe et al. arrived at similar conclusions regarding infanticides as the product of inherently evil males, specifically targetting unweaned infants as lactation stops female fertility. But their conclusions are based on very few cases given the duration considered, and do not check out statistically.
A 10:8 male-to-female attackers ratio over merely 35 individuals does not support his hypothesis, which should see most attacks being made by males on young children of other groups. And not only would it not explain in-group infanticides but they directly contradict the notion of reproductive opportunity killings, because a male chimpanzee has no means to know whether a child in his own group is his own.
This and other such flimsy propositions, as well as their ideological underpinnings—Wrangham and co’s whole thesis looking explicitly to make a good excuse for wars which would be natural and instinctive—are thoroughly highlighted and destroyed with statistical facts and sound arguments, in Brian Ferguson’s work, which contributed more than anyone else to the cause of truth, common sense and animal wellfare.
Intergroup killings of weaned individuals during the two intense conflicts at Gombe and Ngogo produce a rate of 1.7 killings per year whereas the other 416 observation years calculate to .03 per year, so Gombe and Ngogo must be considered outliers, and warrant non-adaptive, non-evolutionary explanation.
Thus most killings were extremely concentrated and removing these outliers restores a much more balanced image, although as we saw before, it is certain that all sites bar none are disturbed, because nearly (hence, not all) all show diseases, a direct evidence of a denatured or very unbalanced diet. The articles acknowledge that no simple explanatory model fits the data. Violence level seems to simply increase indiscriminately at some point, in some places, for some reason, regardless of group size or proximity to other groups.
But there is a very straight-forward explanation: We have been feeding literal shit to animals from 1975 onward, in most observation sites, or at least those featuring murders. This much was confessed by none other than Jane Goodall herself, the fact that killings only started She started “limited human-controlled feeding” of bananas, which significantly changed their behavior from pacific and caring to aggressive, competitive, opportunistically violent as related by Robert Sussmann.
From then on males started being described as fighting over females like bulls. Ferguson correctly incriminates disturbances in the environment, including feedings, but in the article he does agree that no one form of disturbance or model of explanation suffices to explain the form killings take14.
Indeed population density and contact with humans only act as triggers. In our experience territorial animals, if fed raw, fare very well in confined spaces despite their reluctance to intermingle: they segregate and eat on their own. The only form of perturbation that can wreak havoc on the psychological balance of chimpanzees to this point, is molecular perturbation, denatured food.
But placed in dietary conditions as appalling as Neolithic or modern humans, given our genetic proximity one should very much expect the same behaviors: tolerating and seeking bigger groups, tribal conflicts, diseases… and excessive breeding.
Instead of settling on this most parsimonious hypothesis (explaining also the variation in war-like tendencies from group to group and erratic nature of psychological pathologies), proponents of the intrinsic evilness focus on inconclusive studies and disregard the statistical variation.
Constantly news media relate stories of drunk animals, with the obvious intent to clear our conscience, ascribing our addiction to alcohol not to people being utter degenerates (who would survive a week in the wild) but accusing nature itself of having made us potential drunkards. There could not be a better copout strategy than finding drunk animals.
Even if half those cases are fake and the remaining highly misleading, we will show it. It wouldn’t even warrant my time writing this, if a number of scientists too had not been called to accredit those myths.
The problem is similar to that of drugs or other natural psychogenic substances. It can be summed up in 3 points:
Does it exist in wild nature in a concentration great enough to produce any physiological effect ?
If so, are animals instinctively guarded against too high a concentration ?
Can they acquire a preference for it, if so in what conditions ?
Very weak concentrations of alcohol, compared to even beers, arise as a natural product of sugar fermentation in fruits, nectar or sap. But science is definitive on this topic: the quantity is not enough to be a danger. Obviously that will depend on several factors, not just dietary availability (with primates, the amount of fruits rotting on the ground), but the composition of said fermented/alcoholic food in the animal’s diet, and its metabolic rate. By virtue of natural selection, animals routinely eating a big proportion of sugar (especially if susceptible to ferment) will have a bigger liver relative to their size, and higher levels of Alcohol Dehydrogenases (ADH), the enzymes responsible for breaking down ethanol.
And the physiological tolerance to alcohol concentration in our blood also differs, so that only experimental results can decide what does or not cause drunkenness. And observation in relative health rainforests does show that the species consuming high amounts of fermented nectar or yeast do exist but show no sign of intoxication. Similarly inducing alcohol dependence on ape has proven very difficult: they do not like it, neither the taste nor the effect it has on them, and only drink it when forced by circumstances.
One can theorize without too much difficulty, that exposition of the cocktail of thousands of new chemical compounds each cooking recipes magnifies reward circuits a hundred times, creating evolutionary counter-intuitive though a powerful dependency neurologically identical to that of drugs. Unsurprisingly this finding raised much controversy, as it amounts to calling nearly everyone liking sweets a druggie. Yet people very high on industrial sugar match the description to a T. In the same way, the personality of people are addicted to tobacco doesn’t seem altered, but only as long as they can smoke… as they can get a fix.
Cooking as a whole appears much harder to withdraw from than mere just white sugar (or saccharine). It may be essentially socio-psychological—arguably ?—but to master the will to forgo all recipes and follow one’s instinct is more difficult than to stop drinking or smoking, one paar with cocaine addiction. As for the preference for natural sources of alcohol in the wild, no study was ever conducted.
Studies of inborn preference of alcohol in the wild for higher vertebrates are absent. We do have a study about chimpanzees in the wild but with proper alcohol, and not in a form we could possibly ever evolve to regulate. Fermented fruits lie on the ground every now and then. Fermented sap on the other hand lies outside the range of what could occur in nature, and requires human artifices to brew. Chimpanzees drink sap from the tree, never brewing for a minute, let alone a week. A minute of reasoning should suffice to realize that evolution was in no way required to guard us against fermented sap impossible to find in nature.
As all forms of processing brewing is likely to bypass natural regulations. So to assume from the behavior of “wild” chimps eating who-knows-what that the common ancestors of all primates developed an inborn tendency for alcoholism goes beyond stupidity, into the realm of willful deceit of public opinion.
Assuming an uncontrolled natural disposition for alcohol, for megafauna (animals our size or bigger) to get drunk one would require enormous amount of fermented fruits at the same place and time incompatible with any notion of instinctive regulation, so that an automatic vomitive reflex would kick in before the alcohol level could rise in any capacity. It is a fact that natural food almost never reaches beer-levels of ethanol, much less degrees higher than 3%.
So in any case to argue an hypothetical taste for a 1 to 1.5% degree in food still wouldn’t account for humans’ addiction or taste to the likes of cider (3%) beer (4.2%) whine (5%) sake (15%) or whisky (40%). The tolerance to a certain substance selected by evolution has to reflect the conditions found in our environment, which never included anything higher than 2% at the very worst in any appreciable quantity. Therefore no evolutionary argument can explain a taste for alcohol beverage.
Only the paradoxical physiological state induced by cooking, plus the early annihilation of our dietary instinct could, plus a strong cultural psycho-cultural conditioning to do away with the inborn aversion every child experiences. That is why captive primates have been seen liking some level of alcohol
As ethanol concentration increased, drinking (the alcoholized) water increased, and total water decreased. Net ethanol intake remained relatively constant, and could lead to notable blood ethanol levels. However the diet consisted of “regular monkey pellets” supplemented with fruit at 1340 hours and a slice of bread. If we consider the digestive discomfort and heavy sleep cooked food cause brings, the alleged digestive and appetizing effect of alcohol at low levels, and the capacity of non-human primates to offset more immediate reactions in order to repeat a positive outcome they experienced, instead of seeking tipsiness those monkeys must have sought the momentaneous relief from permanent intestinal discomfort and the speeding up the expulsion of denatured molecules—pooping—that some amount of alcohol provided.
The precise regulation in ethanol consumption observed might allude to a certain dose of fermentation (indicated by ethanol) being not just benign but desirable, due to us having adapted to it. Some even spontaneously form in our guts. The monkeys may have just sought to compensate for the lack of it in their lab diet. Typically, children learn to break open aversion barriers to coffee and alcohol by masking their taste with sugar, preferring sweet beers or cider to wine. Sugar + ethanol might as well explain why people start drinking, the combination unconsciously reminding their body of fermented fruits.
drunken birds run over by a car
Now on to birds: it has been argued in recent years, that birds also frequently get drunk on fermented fruits and die. Let us examine these claims.
Some birds have a very specialized diet, consisting mostly if not totally on fruits. So like the tree shrews, they evolved to ingest, digest and excrete in a matter of an hour up to the equivalent of a man of 70 kg eating 14 kg at once, not even throughout the day. Their metabolism, as per the requirements of constant flight as well as thermogenesis (in both cases the smaller the less efficient energy-wise) is—as general rule—much faster than mammals’, and the smaller the faster. Hence for a given feeding/digestion time they will be in contact with a lot more alcohol per gram of body mass than would frugivores among mammals. Then those levels quickly dwindle as the ethanol (then more importantly, its toxic subproduct acetaldehyde) breaks down due to enzymes.
But do birds always cope with their diet ? Natural selection would say “Yes of course duh” but people keep reporting slews of wild birds dropping from the sky, running into windows and cars, singing on the street in groups oblivious to humans, unable to fly off. Seemingly completely hammered. And people love to report it, incriminating fermenting berries.
See the New York Times, in 2018, giving up this self-comforting popular telltale of drunken birds. Scientists have been debating for decades by now, owing to the difficulty of analyzing the lots of dead or alleged drunken birds by forensic pathologists, in order to ascertain common causes of possibly unrelated accidents. One difficulty lies in the variety of species involved with variation in the metabolism of the alcohol-breaking enzyme ADH reaching up to 23 times, forbidding general conclusions as to what is within physiological range or not.
That behavior alone, ethanol-induced stupor or not, doesn’t seem normal at all. Both populations and needs should be tailored to the quantities and diversity of food, so that animals shouldn’t feel any such rush, not to the point of killing themselves, when less ripe fruits exist. If for some reason in tens or hundreds of thousands of years that very parameter (availability of ripe berries) did not change, then birds would adapt to eat less ripe ones. And coevolution—birds spreading seeds through defecation—would favor species ripening earlier or longer.
The fact it didn’t happen, means that there used to be enough food so that such selection pressure couldn’t increase.
In some cases some birds fall back on back on less appealing, too fermented berries, pushing the limit, with deadly consequences. But how many of them is highly debatable and was debated to death, as many such fallen birds have been analyzed, depending on the species some autopsies demonstrate ethanol intoxication, and some don’t. It doesn’t take much to realize that in the past, a mere 50 years ago, urbanization in the US wasn’t nearly as extensive and intensive as today. There were a lot more trees, more diverse too, the farther back in time we go. So… environmental degradation, as usual. The simplest cause is merely overeating, due to the extreme rarity of ripe fruits in the urbanized, semi-urbanized or agricultural lands: that much was concluded by sound minds. Intoxication might very well occur as a consequence of overeating and excess sugar level, but no one ever showed that (wild) birds prefer fermented berries so in no way did these happenings ever help condone our alcoholism. Intoxication is simply always a consequence of starvation and overeating.
We find confirmations in the latest state of cellular biology and genetics, that viral processes are in reality in most cases, under the control of the body, and that viruses, exosomes (and prions) are all variations of an universal mailing system shared by all of nature for billions of years.
Among viruses, those that interest us are the “enveloping viruses” because they cover their protein capsid (if they have one) and their genetic material by one (or more) lipid bilayer. They can derive either from nuclear membranes like the envelopes of Herpesviridae or from plasma membranes for Orthomyxoviridae. The genetic material of these viruses can be of any type: RNA, DNA, single or double stranded.
Exosomes in theory, as vesicles, are not related despite their similarity because unlike viruses they do not reproduce. However, contemporary virology has moved away from this strict definition by its wide use of the terms non-infectious and defective virus. Therefore, extracellular vesicles generated by retrovirus-infected cells that carry viral proteins and even fragments of viral genomes essentially fall into this category of “non-infectious viruses”.
They are surrounded by a lipid membrane that also contains cell membrane proteins. Like those of viral envelopes, these proteins can determine adhesion to the plasma membrane of specific target cells. In addition to DNA polymerase activity, tumor vesicles also showed endogenous reverse transcriptase activity more commonly associated with viruses.
Despite the important differences in membrane morphology, recent research has revealed a variety of similar mechanisms in bacteria, pointing to an identity of form and function of vesicular formations between the three domains of life (bacteria, archaea and eukaryotes or cells with nuclei including animal, plant or fungal organisms, unicellular or multicellular). I will only mention them here in the spirit of completeness.
a: bacterium, b: eukaryote, c and d: archaea
Bacteria display a layer of peptidoglycan on their inner membrane, a rigid substance giving very particular conditions for the budding of vesicles on the surface. The vast majority of bacteria also have a layer of external lipopolysaccharide (LPS) and underneath a little peptidoglycan located in the periplasmic space. They are generally referred to as Gram-negative or bidermic bacteria. The composition of Gram-positive and Gram-negative varies15, but several types of extracellular vesicles have been described.
Vesicles are associated with long filamentous structures connecting cells in all three domains of life, including nanotubes, tunnelling nanotubes and nanopods. The most studied being formed by budding from the LPS-containing outer membrane (OM) and containing mainly periplasmic components, however some formed from the outer and inner membranes of didermatid bacteria have been identified in several species, containing elements of the whole bacterium. Finally, vesicles have been observed in Gram+ bacteria with thick envelopes, crossing the thickness.
They share basically the same characteristics of eukaryotic vesicles and viruses: resistance to enzymatic attack, molecular profiling allowing a selective delivery of their payload at long distance with high specificity toward a target cell while passing immune cells’ checks. They can also help bacterial colonization by selectively killing or promoting the growth of other bacterial species, by exchanging genetic material (“transformasome”). And as in the animal case, increased vesiculation may aid in removal of toxic by-products after exposure to stress.
These nanotubes (membranous structures 50-200 nm in diameter and up to several cell diameters in length) often contain membrane-surrounded vesicle arrays, suggesting that they may be involved in vesicle production, particularly by some cancer cells
Different morphologies of extracellular vesicles
But we know a lot more about vesicles. Due to the small size and heterogeneity of vesicles, their detection and classification is difficult. Although different types of vesicles have been identified, widely used terms such as “exosomes” “ectosomes” and “microparticles” are often inconsistent, especially in the older literature. In addition, the extent to which the various morphologies contribute to the processes studied is largely unknown. For practical reasons, the various terms are to be considered here roughly interchangeable. Electron microscopy remains the gold standard technique for determining vesicle morphology and size.
Just like viruses, exosomes can cross biological barriers (such as the blood-brain barrier) and penetrate cells with a high degree of specificity. Hence their significant interest as drug delivery agents and diagnostic biomarkers.
In general, their diameter does not exceed 120 nm, which is smaller than the maximum theoretical resolution of a conventional optical microscope. The fate and interactions of exosomes inside cells are therefore difficult to study, which limits investigation in the field, although newer techniques (PALM and Dstorm) overcame the limit of light diffraction and allow the examination of exosomes and their contents down to the molecule level.
Populations of extracellular vesicles are very heterogeneous, even in pure cell culture, each cell type being able to produce different types. However some seem produced exclusively by certain cells. To further complicate matters, the content of extracellular vesicles varies depending on the source and the original isolation or enrichment technique. Their method of budding or creation from cell membranes vary a lot, but much more is known about it than about the loading of nucleic acids.
In one study, it was shown that double-stranded DNA would rather be adsorbed on the outside of vesicles and large vesicles carry most of the tumor double-stranded DNA circulating in the plasma of prostate cancer patients. Because of their oncogenic potential much data on extracellular vesicles in the context of tumors have been collected, but this also reflects the fact that the spread or functions of endogenous exosomes in vivo or data on adult tissues under healthy i.e. homeostatic equilibrium conditions are pretty much unknown, in comparison.
The existence of sorting mechanisms is assumed but not known. Several sequence motifs highly enriched in exosome-associated miRNAs (an important class with a ubiquitous role as a negative regulator of gene expression), compared to cellular miRNAs, have been determined, along with a specific protein binding to these “EXOmotifs”, thus triggering RNA loading.
Sequence motifs on messenger RNAs lead to vesicle enrichment via a mechanism involving interactions between mRNAs and microRNAs, sometimes involving specific transcription factors binding degenerate consensus sequences in the upstream untranslated region (UTR) of the RNA.
Computer simulation of spontaneous membrane vascularization
Conceptually, virus budding can be divided into two steps:
membrane deformation, when the membrane is “wrapped” around the assembling virion, then membrane fission, when the neck of the bud is severed.
The structural proteins of enveloped viruses generally bind to membranes and form spherical or helical assemblies. Thus, assembly and budding are often inextricably linked processes. These fusion proteins undergo dramatic conformational changes, converting the free energy released by fusion protein folding into energy that is used to fuse viral and cellular membranes. Exosomes share the same mechanism and in some cases the same cellular pathways (such as the ESCRT pathway) have been observed.
Budding is usually tightly coupled to virion assembly, so most viruses use their structural proteins to recruit the ESCRT pathway. They can also use host factors, for example in the GAG polyprotein (the main structural element of retroviruses). In HIV-1, the energy released is not required for Gag polymerization but for detaching nascent virions from the plasma membrane. Subsequent studies identified two different short peptide motifs in p6 Gag that contributed to the efficiency of HIV-1 budding.
In parallel, others have identified distinct short peptide motifs in the structural proteins of other viruses, called “late assembly domains” of which at least five distinct but interchangeable classes have subsequently been identified, often leading to the identification of analogous motifs within cellular proteins, recruiting ESCRT factors, an interaction verified experimentally. Funny system where thieves and bankers agree on a common system of semaphores to keep each other in sight.
Exosomes and viruses share, in addition to budding, probably the mechanisms of specific cargo packaging and membrane budding for cell release. Most surprisingly, different viruses with very different evolutionary pathways seem to converge in their use of the endocytic pathway for entry and exit of their host cells.
Similarly, the hepatitis C virus is known to incorporate its entire RNA genome into exosomes without surface proteins, leaving them “infectious”. Most of these data on the function of extracellular vesicles such as exosomes, however, are collected from transformed cancer cells and depend on the use of a heterogeneous population of vesicles purified from supernatant or liquid cell cultures. Therefore, the spread of endogenous exosomes in vivo is largely unknown, and data on their biogenesis and role in normal development of tissues and adult tissues under homeostatic conditions are clearly lagging and understanding their transfer and fate in recipient cells is crucial.
Eukaryotic exosomes contain large amounts of RNA of all kinds and transfer these to target cells. Moreover this RNA content is highly specific and sometimes enriched several thousand times relative to their host cells even between different subpopulations of the exosomes. RNAs isolated from apoptotic vesicles (formed after programmed cell death), membrane vesicles and exosomes have very different profiles and effects on target cells
It can only imply a purposeful packaging, and reflect an active process of communication to other surrounding cells informing about the causes and conditions of apoptotic cell death.
Although there is intact mRNA and long non-coding RNA present in EVs, most RNAs are fragmented or small in size. As mentioned above, these RNAs encapsulated in vesicles can have a profound impact on recipient cells, transferring between different cell types a signal causing transient transformation of recipient cells e.g. leading to the production of new proteins (case of mRNA transfer), or regulation of gene expression in the case of miRNAs.
That means they are actively packaged with a purpose, and reflect an active process of communication to other surrounding cells informing about the causes and conditions of apoptotic cell death.
The viral process is only one aspect of intercellular communication, including between different organisms, although in ecological ways that are not yet understood. Through microRNAs, some viruses can modulate cellular processes as diverse as immune evasion, apoptosis, proliferation, and even viral infectivity. These viral miRNAs in conjunction with endogenous miRNAs are thought to play a role in modulating the expression of target genes in recipient cells. In addition to RNAs, infected cells may also excrete specific viral proteins via exosomes, thereby modulating cellular processes in surrounding cells (cytokines, interferons).
For example: exosomes released from HTLV-1 (human T-lymphotropic virus 1) infected cells contain not only viral mRNA transcripts, such as those of Tax, HBZ and Env, but also the biologically active trans-activator protein Tax. In addition, the HTLV-1 Tax protein was shown in exosomes isolated from cerebrospinal fluid of patients with HTLV-1-associated tropical spastic paraparesis myelopathy, suggesting that HTLV-1 may modulate its microenvironment by selective secretion of specific viral cargoes.
Substantial evidence indicates that many different types of pathogens, including bacteria, viruses and protozoa (and prions, see below) can “exploit” the exosomal pathway, for secretion or movement. These “parasites” can also influence exported cellular products. A body of evidence indicates that exosomes in virus-infected cells can induce processes as diverse as immune evasion, apoptosis, angiogenesis, proliferation, trans-cellular spread and cytokine modulation. The molecular details of how these processes are triggered are poorly understood, and they differ between tissues, cell states, viruses…
Another well-known mechanism by which some viruses can evade immune responses is by down-regulating viral lytic gene expression and persisting in infected cells in a latent state, where the absence of viral antigen in “infected” cells means that the immune response cannot be triggered, but also simply that the virus is “paused.
For example, herpes simplex type 1 (HSV-1) replicates in mucosal epithelial cells during primary infection and then enters the sensory neurons where it establishes a lifetime latency. During the latent state, although no viral protein is expressed, numerous miRNAs (viral microRNAs) have been detected, and some of these miRNAs appear to play a central role in suppressing viral gene expression and maintaining latency. And transmit antiviral factors, to the same effect.
There is some evidence that EVs, although less efficient than virions themselves, can transfer cytosolic proteins involved in antiviral responses, such as APOBEC3G and cGAMP (33-36) to recipient cells. In addition to miRNAs, extracellular vesicles also contain a wide variety of other small noncoding RNAs, such as fragments of protein-coding regions and repeat sequences, which could also act as regulatory RNAs by influencing gene expression.
In some viral infections, such as hepatitis B and herpes as well, non-infectious subviral particles are released into the serum, in some cases without viral capsid or viral DNA, often at levels 1000-fold higher than mature infectious particles. One hypothesis is that these subviruses would lure the immune system. In addition to immune modulation, exosomes released from some virus-infected cells may promote viral infection and spread as discussed with HIV-1.
In a similar vein, T cells can produce vesicles containing the HIV CD4 receptor, allowing them to bind to viral particles as antibodies would, thereby decreasing the number of virions that would otherwise affect CD4+ T cells. Thus, the cells are able to selectively exchange antigens with each other, certainly to direct the process of clonal selection in secondary lymphoid organs.
Conversely, in vivo, exosomes interact with viruses and each other directly or via modulation of host responses thus participating in a “war and peace” between viruses and the host. In 2014, exosomes derived from infected cells containing Tax proteins and pro-inflammatory factors, as well as viral mRNA transcripts including Tax, HBZ and Env (viral envelope protein) were found. In addition, giving these Tax-expressing exosomes to other cells improved their cell survival to Fas antibody treatment, indicating induction of NF-kB and activation of AKT (2 anti-apoptotic factors). Another study found similar processes for HIV. When transferred via exosomes, TAR RNA can increase the population of susceptible target cells. Inside EV target cells, full-length TAR RNA is transformed into miRNAs, which quench the mRNA encoding the Bcl-2 interacting protein (involved in induction of apoptosis).
The Trojan exosome hypothesis proposed that HIV evolved to use the exosomal system (more precisely ESCRT and Rab GTPases), in the absence of a receptor-based system, by transferring the virus to closely associated uninfected T cells via exosomes… Furthermore, its infectivity is reduced in the absence of exosomes. But given other viruses can do this, we would argue it is not a feature of viruses themselves but a pathway cells dispose of, to broadcast a message.
Viruses of several different species commonly were found to travel in the same exosomes throughout the body, “collaborating” in order to improve the collective fitness of the viral population, and thus were (contrary to what was thought) in physical proximity, creating the perfect conditions for increased horizontal exchange.
The mentioned article still talks about “exploitation” of vesicular trafficking and “genetic cooperation” between viral species, as if it were a criminal alliance with a will to do harm. But viruses have no more will than they have metabolism, they are only molecular robots, and they are entirely passive.
In particular the content of HIV is very limited, known at the tip of one’s fingers. One could well compare it to a computer whose program in assembler has so well elaborated to take part of the instruction set of the CISC processor (for “Complex Instruction Set Complex”, hard to imagine more “complex than an animal cell!). And to keep this extreme syntony, in spite of this same enormous mutation rate which allows it to escape the immune system and vaccine attempts.
So there is no reason why the interaction between viral proteins of different species should produce anything other than disorder: Putting a helical gear there instead of a normal straight road is not known to improve a mechanism, and a cell is several hundred times more complex than a machine, in fact scientists compare it more to a whole city than to a factory. What kind of miracle of improbability would it take for virologists to question their view?
This proximity and this traffic, is one more proof of the arranged and programmed nature of the viral phenomenon, in concert with the cellular dynamics. And not the blind product of a mutative process still much more chaotic on average than our cells. Typically, influenza, HIV and coronaviruses mutate several dozen times. And yet the information necessary for proper functioning is maintained. So we invoke, as always… the sacrosanct “natural selection”.
But since it is an article of faith without the slightest statistical justification or mathematical model (the complete “life” cycle involving the whole cell, thus exceeding our computers by several millions or billions) we don’t need to ask ourselves any questions, and we will put forward the same argument no matter what the facts are that go against the grain and the enormity of the claim. This defines a dogma, a religion.
Other functional studies of exosomes released from infected cells have shown that infected cells release not only infectious virions, which are capable of spreading infection, but also a variety of non-replicative particles, which are difficult to classify, as they could be considered either as defective viral particles or vesicles containing viral elements, such as viral proteins and viral regulatory double-stranded DNA.
These non-replicating particles typically include cytidine deaminase, degrading retroviral RNA by random mutation, inhibiting viral replication. Exosomes containing host miRNAs produced by virus-resistant cells can confer resistance to other cells. This has been demonstrated for trophoblasts, which are largely resistant to infection by various viruses, including HIV, likely contributing to fetal protection in vivo. Exosomes produced by these cells in vitro carry host miRNAs and deliver them to virus-susceptible cells, making them resistant to virus infection.
One can see the analogy with the aforementioned “defective viruses” indistinguishable from exosomes, which confer resistance AND at the same time reduce the immune response. They are two sides of the same phenomenon.
In humans, prion disease occurs in sporadic, familial and acquired etiologies. However, all forms of the disease are transmissible, with possible routes of infection through dietary exposure, medical procedures and blood transfusion. The isoform of the normal prion protein, PrP C, is encoded by PRNP and is expressed in all tissues of the human body, with the highest levels of expression observed in central nervous system and brain tissues.
PrPC and PrPSc have been isolated in association with exosomes, and PrPSc-containing exosomes were infectious in both animal and human cell bioassays. Alzheimer’s disease (AD) is the most common form of dementia in humans and is pathologically characterized by the extracellular deposition of insoluble amyloid plaques consisting of β-amyloid peptide (Aβ), a 39-43 amino acid peptide produced by proteolytic cleavage of the amyloid precursor protein. APP is cleaved from Aβ, left intact as a membrane associated fragment (β-CTF). These fragments accumulate intracellularly in multivesicular bodies and are incorporated into exosomes and released into the extracellular environment.
The identification of Aβ in association with exosomes is an important finding, especially since other exosomal proteins such as Alix and Flotillin-1 have been shown to accumulate in the brain plaques of Alzheimer’s patients. Exosomes could also provide an explanation for the transport of equally toxic Aβ and APP-CTF around the body to the brain, where they contribute to amyloid deposition. Underlying causes of neurodegenerative diseases are the folding and aggregation of specific proteins, such as amyloid β peptide (Aβ) in AD, scrapie associated prion protein (PrPSc) in prion disease, α-synuclein in PD, and superoxide dismutase 1 (oxidoreductase catalyzing the dismutation of superoxide anions O2–, involved in the neutralization of oxidative stress) in amyotrophic lateral sclerosis.
And all of these enzymes seem to be moving in the same direction: cleaning up, removing metabolic waste. Even if the theory still lacks precision, it seems clear that prion and prion-like diseases are neither “infections” or diseases, but essential vital clean processes of eliminating denatured molecules, AGEs in particular. Alongside or at the same time as prions, the main proteins associated with these neurodegenerative disorders are found to colocalize on/in exosomes, containing binding sites for copper, zinc, iron and manganese. All of these have an obvious role in what would normally be detoxification: a dozen zinc transporters, ferroportin, transferrins, serotransferrin, several metalloreductases, ferritins, aconitate hydratase (sequester iron), ferroxidase, for iron.
We have already seen the role of AGEs and ALEs in disorders as varied as cancer, diabetes, aging. They are found colocalizing with AGEs receptors (RAGEs) in exosomes with Alzheimer’s disease(Vlassara and Patterson). RAGEs also bind the amyloid substance responsible for plaques, stimulating its production in the brain and regulating its influx across the blood-brain barrier. Non-enzymatic glycation of proteins has been reported to stimulate protein aggregation and amyloid deposition. They promote senile plaque formation at least via tau hyper-phosphorylation, synaptic dysfunction and neuronal death.
Prions must constitute an extensive set of proteins with several functions important to the brain (mostly but not only), one being the sequestration of denatured molecules. However, the formation of plaque, linked to age and therefore to the level of intoxication, is not their property: rather, it is the consequence of a rate of intoxication completely unforeseen by nature and reaching such a peak in old people that the body is overwhelmed.
Certain cells in the central nervous system, expectedly, appear as specialized cleaner team, clearing “malformed” protein aggregates, whether innately toxic or acting as a buffer between maintenance cells and really toxic aggregates, neutralizing them the way antibodies surround antigens.
The body does not treat all “misfolded” proteins in the same way some being toxic some not, so obviously will be waste products, while others (amyloids and other β-sheets in particular) as normal tools against reaction to oxidative stress, that because of the excessive increase due to the unnatural levels of AGEs we have confused for the diseases’ cause. Both AGEs and amyloid substances accumulate with aging, so we believe at least some of them constitute natural molecular defenses against normal metabolic waste, part of which coincide with the products of cooking symptoms in quality, although not in quantity, hence the excess observed to the point that patients’ brain end up as porous as fine gruyere. Other misfolded proteins likely fall into the waste category themselves, and come from the inside or diet.
So we have seen infectious exosomes that are not viruses, and viruses that are not infectious, “not quite infectious”. Since the purpose of the phenomenon is not negative for the organism (beyond the individual cells), is it logical to always consider these viruses as “foreign” when we observe a whole continuum between self and non-self?
As far as the DNA content in eukaryotic extracellular vesicles is concerned, less is known than the RNA content. Single-stranded, mitochondrial, plasmid, and double-stranded DNA have been observed. The DNA seems to be present in the form of small fragments (about 10 to 20 kb) covering the whole genome including mitochondria without apparent order for the moment. However, it is important to note that some EV-associated DNA fragments contain entire genes with promoter and terminator regions. This DNA can be transported from cell to cell by endocytosis or fusion and this transfer can affect the transcriptional pattern of recipient cells, inducing both up- and down-regulation of many genes.
Finally, not only RNA from enveloped retroviruses but also non-enveloped DNA viruses, such as herpesviruses, export their products and genetic material in the same way. Many aspects of this mechanism remain to be clarified.
Just as the majority of viral “infections” are said to be “frustrated”, the detection of the “pathological” PrPSc isoform is not a sign of a disease, there would be “thresholds”, and an excess of normal forms would be able to trigger the conversion.
The consequent increase in resistance to apoptosis is said to allow the cell to produce a virus for a longer period of time, thus facilitating the spread of HIV. This is true, but the process is not automatic or a priori undergone by the cells and one can decently ask the question: why in the same package, incorporate virions at the same time as deaminases decreasing the efficiency of the viral load? If we exclude this disturbing fact, the official theory of the pathogen already holds little when we consider that HIV for example would contain only 10 genes…
What an incredible compaction of information, if viruses can with so little orchestrate a whole set of cellular and inter-cellular reactions, and program exosomes to target specific cells or organs. Well, essentially the same characteristics must be attributed to a simple deformed protein (the prion), having evolved according to the official theory to take control to the point of pushing distant organs to send their own quota of proteins to the brain.
We urgently need to change our perspective and see it as simply an information exchange protocol, just like sex hairs, more classical viruses, tunnels, circulating RNAs and maybe something else yet to be discovered. From my search of the literature, no one has thought of tracking the movement of a single AGE molecule, for example cells in the gut wall, from a nutrient solution that mimics the natural nutrient concentrations from a meal. However, this would be technically feasible16.
It is important to be able to show directly a correlation between reciprocal behavior in case of “infection” by different viral agents, and the treatment of this xenobiotic load and/or the packaging in the exosomes themselves. Similarly, tracking a prion or Aβ protein as it travels through the body would provide crucial information about how cells process the signal and pass it on, as on potentially several meters of transit there is a chance that the exosome carrying them will be contacted or even captured by cells for which they are not intended.
Isolated bird populations of known origins revealed a mechanism) that compensate for the low level of MHC diversity in genetically depauperate population, by reshuffling exons while maintening the exclusion from point blank SNPs, thus preserving functionality.
Given that the process takes at minima a few tens of thousands of years (starting with less than a hundred strong founder population), most likely only in the few rare individuals, the frequency of the mechanism per site and generation must still be extremely rare… an order of magnitude higher than the expected chance mutation, while significant, is still very low. This could explain the difficulty to see this phenomena in laboratory or other controlled conditions, yet should be added to the three aforementioned classical mechanisms.
The difference being, that this scenario does not involve natural selection, pathogene-mediated or otherwise, with its slew of unfits dying: While individuals with a renewed MHC repertoire could boast a survival advantage the latter is likely not significant, because the phenomena takes place in the span of millenia, not decades or centuries. Strenghtening the absence of experimental evidence for natural selection against pathogens.
Explaining this incredible natural polymorphism demands quite a lot intellectual gymnastics, to explain how a supposed strong selection (which pathogens should cause) can coexist with an absence of selective sweeps (the eventual fixation of a beneficial mutation) and thus the maintenance of infrequent alleles, in less than 1% of the population.
The ruling opinion dictates that pathogens mutate constantly so that heterozygotes always have an advantage (heterozygote advantage), alleles never have the time to be discarded completely before recovering again some protective value (fluctuating selection) and rarity would hold an intrinsic edge for minor haplotypes as microbes would wouldn’t statistically be compelled to adapt to unfrequent alleles, thus keeping the latter both relevant and rare.
It all sounds fine and dandy, but proving this model or the prevalence of any one of these mechanisms is impossible to attest or prove, both experimentally and theoretically, thus it it is not a scientific theory but make-believe to maintain the myth of diversity, natural selection and the germ theory. And one should not conclude that cells would somehow keep a record in order to strike something like a “golden mean” in diversity, this gene conversion actually erased a number of haplotypes in local populations, their number only rising at the metapopulation level.
Several arguments in this case serve to discredit against the pathogen-driven selection being behind the usually high number of polymorphisms in the MHC.
The fact this increase here took 75 000 years at minima, implies the initial low MHC diversity in this bird population must not have prompted a strong selection pressure or caused significant mortality.
Non-synonymous mutations did not exceed synonymous/neutral ones, a signature usually interpreted as the absence of positive selection. Yes hitchiking could explain that, however
this phenomena only shuffled the existing, with no creation of new functional alleles, so that recombinant haplotypes could hardly constitute new effective defenses, which would quickly loose their edge anyway as supposedly pathogens never cease to evolve, and much faster than animals too.
75 000 years should be more than enough to overcome a very limited repertoire and we ought to see balancing selection at play, with diversity increasing both globally and locally.
What we see instead is a very slow coalescing of an optimal number of semaphores with which to perfect the dialog with the local microbes and parasites. I claim that the best fitting hypothesis for the genetic facts we observe in the MHC, are best explained by vertebrates sharing from time immemorial a symbiotic relationships with all microbes and all parasites, commanding them due to the exponential gap in complexity and adaptive capacity in our immune system.
The principle being, that it happens only as a consequence of pollution, or metabolic deficiency leading to waste accumulation and immune functions (meant to keep exogenetic cleaners honest) failing due to old age, signaling to predators and parasites alike which animals to single out, thus keeping populations in optimal health. Messed-up genetics from artificial or accidental excessive inbreeding may also be expected to lead to metabolic imbalance thus eliciting the appearance of pathogen-driven selection. Such models can not inform natural situations: these animals are not only very homozygous, but very degenerate too.
Assuming no human disturbance or cooking, no evidence of a genuinely adverse relationship has been seen with a natural microbe or parasite, in a natural situation, in Europe or Africa, on people or domestic animals, no any issue with local or exotic food as genetically remote and alien-looking as can be, pointing at the existence of an universal molecular order as old as life itself, explaning for instance how primate palates can enjoy and digest deep sea food, despite their ancestors possibly possibly not tasting them since the first batracian ventured on land.
So I supopse that in absence of environmental or dietary pollution mortal infections should not follow exposure to new “pathogens”, when it comes to mammals, reptiles if not fish too. The concept of natural selection (in the strict sense of death and survival of the fittest) by pathogens in the case of higher vertebrates (and a small number of other animals), ultimately derives from the same pasteurian mindset as germ theory itself. We learnt to see only fighting and adversity, when nature has always been all about balance all along. Maybe it is time we accept to revise our deep-seated presuppositions, and see this balance not just as an emergent property of complex ecological systems (species endangering their own long time survival weeding themselves out eventually) as modern ecology theorizes, but as a truly primary force ruling shaping genetics and molecular biology with undercurrents actually more fundamental than Darwinian dynamics.
When I’m told that 50 percent of dogs die of cancer, there must be an explanation for that. Nature has predisposed the dog to feed on raw meat, by tearing up other animals. To-day the dog feeds almost exclusively on mixed bread and cooked meat.
Country folk spend fourteen hours a day in the fresh air. Yet by the age of forty-five they’re old, and the mortality amongst them is enormous. That’s the result of an error in their diet. They eat only cooked foods. Everything that lives on earth feeds on living materials. The fact that man subjects his foodstuffs to a physico-chemical process explains the so-called “maladies of civilization”. If the average term of life is at present increasing, that’s because people are again finding room for a naturistic diet. It’s a revolution. That a fatty substance extracted from coal has the same value as olive-oil, that l don’t believe at all! It’s not impossible that one of the causes of cancer lies in the harmfulness of cooked foods. We give our body a form of nourishment that in one way or another is debased. At present the origin of cancer is unknown, but it’s possible that the causes that provoke it find a terrain that suits them in incorrectly nourished organisms. We all breathe in the microbes that give rise to colds or tuberculosis, but we’re not all enrheumed or tuberculous.
Nature, in creating a being, gives it all it needs to live. If it cannot live, that’s either because it’s attacked from without or because its inner resistance has weakened. In the case of man, it’s usually the second eventuality that has made him vulnerable
The consumption of meat is reduced the moment the market presents a greater choice of vegetables, and in proportion as each man can afford the luxury of the first fruits. I suppose man became carnivorous because, during the Ice Age, circumstances compelled him. They also prompted him to have his food cooked, a habit which, as one knows to-day, has harmful consequences. Our peasants never eat any food that hasn’t been cooked and re-cooked, and thus deprived of all its virtues.
At the time when I ate meat, I used to sweat a lot. I used to drink four pots of beer and six bottles of water during a meeting, and I’d succeed in losing nine pounds ! When I became a vegetarian, a mouthful of water from time to time was enough.
When I was a young man, the doctors used to say that a meat diet was indispensable for the formation of bones. This was not true. Unlike peoples who eat polenta, we have bad teeth. It occurs to me that this has something to do with a diet that’s more or less rich in yeast. Nine-tenths of our diet are made up of foods deprived of their biological qualities.
In Sologne, known for its insalubrity and poverty, Abbé Tessier, sent on a mission by the Royal Society of Medicine to investigate ergot rye, known for its abortifacient power, noted: Women are very fertile, although thashey are not regulated before eighteen or twenty years of age: they cease to be so at thirty-six and forty. They have up to fifteen children.
The worthy abbot emphasizes that this precocious inclination (to early love-making) is due to the free communication of boys and girls who guard their flocks together and sleep in the same room, but this nevertheless does not hasten an early puberty in any way.
The children are very healthy and very colorful as the girls are brought up in a difficult exercise, far above that which is necessary for the exercise of their functions, they are adjusted very late. The influence of physical work is underlined here.
[in Brittany, Vieillevigne] Among people of the sex, those who are not employed in rural work are settled from eleven to fourteen years old, those who are are occupied only from fourteen to eighteen years of age… (1787)
If girls who did not work had their periods earlier, it was undoubtedly because their parents had enough to feed them properly (sic!) by doing without their services in the fields.
Among the women of Haute Auvergne, according to Doctor de Brieude:
The constitution of women is sanguine like that of men: their weak and loose fiber makes the difference, because it makes them more humorous. Menstruation does not appear in them until the age of fourteen or fifteen; we find in the countryside where passions are silent longer, girls who have reached their eighteenth year without having seen anything, who nevertheless enjoy good health. When they flow easily, their duration is six days in a person of twenty to thirty years; they are abundant for three days; this evacuation does not end until around forty-eight or fifty years of age… [Brieude, 1782: 300]
High ages at first menstruation are cited for several municipalities in the department of Seine-et-Marne between 1806 and 1815. It is these data which form the last point of this article. The women are also of average size, they are very fertile. Menstruation rarely begins before the age of 17. The menstrual flow which in the girls of Achères does not appear very early continues until the age of 50 and 55, which means that they are fertile for a very long time; It is also not uncommon to see women aged 40 to 45 become mothers, something that is not commonly observed in large cities where menstruation begins at 12 or 13 and disappears between 40 and 45. It follows that it is a real advantage for the population that the menstrual flow does not establish too early. Moreover, we would be very mistaken if we considered early menstruation as the product of a more robust constitution, or a particular development of the genital organs. The opposite is observed every day.
Whilst they are inside we shut them in so that we can have access to the exterior cages to add fresh straw or do maintenance etc. We also give them their first feed or breakfast which we put on the roof of the cage “making” the gorillas work for their breakfast and spreading it out so that everyone gets a fair share. As wild gorillas naturally spend most of the day foraging we give the second feed as another scatter feed, which usually consists of nuts, a specially formulated pellet (which looks a bit like “Cheerios”) and small or chopped items of fruit. We would normally distribute this in the gardens to encourage them out into the open at 12 O’clock.
We used microsatellite genotypes from museum specimens and contemporary samples of Cross River gorillas to infer both the long-term and recent population history. We find that Cross River gorillas diverged from the ancestral western gorilla population ~17,800 years ago (95% HDI: 760, 63,245 years). However, gene flow ceased only ~420 years ago (95% HDI: 200, 16,256 years), followed by a bottleneck beginning ~320 years ago (95% HDI: 200, 2,825 years) that caused a 60-fold decrease in the effective population size of Cross River gorillas. Direct comparison of heterozygosity estimates from museum and contemporary samples suggests a loss of genetic variation over the last 100 years.
The composite history of western gorillas could plausibly be explained by climatic oscillations inducing environmental changes in western equatorial Africa that would have allowed gorilla populations to expand over time but ultimately isolate the Cross River gorillas, which thereafter exhibited a dramatic population size reduction. The recent decrease in the Cross River population is accordingly most likely attributable to increasing anthropogenic pressure over the last several hundred years.
Given that this also coincides with a notable decline in western lowland gorilla Ne, it may be that environmental changes during the Last Glacial Maximum (26,000 to 19,000 years ago), when dry savannah replaced tropical forest over much of the Congo basin (21), triggered a collapse in the western population and complete separation of the two species.
Bwindi mountain gorillas spent on average 15% of feeding time consuming fruit, with monthly variation ranging from 0 to 70%.
This study confirms that gorillas are frugivorous, and only the Virunga mountain gorilla population can be characterized as highly folivorous. Along with other frugivorous great apes, Bwindi mountain gorillas alter their activity patterns in response to varying amounts of fruit in their diet.
In contrast to the less than 1% of foraging time spent on fruit in Virunga mountain gorillas, western lowland gorillas spend approximately 30% of their foraging time consuming fruit, accounting for up to 70% of feeding time during peak fruit season well as within and between years, and within and between sites.
Free-ranging gorillas consume a wide variety of plant species, with 50 to 300 species reported. However, they are very selective, choosing only certain parts of the vegetation at certain times of the year. As an example, only the base and tips of young leaves may be selected, although mature leaves are also eaten. Even captive gorillas are selective feeders, with particular species and plant parts preferred by individuals and groups. Gorillas generally select immature leaves over the mature ones, which usually contain less fiber, more protein, and less secondary components such as tannins. Shoots, flowers, and fruit are also preferred over mature leaves.
The gorillas in Lope Reserve do not appear to select food based on any antinutritional properties (i.e., phenolic compounds), and consume a wide variety of fruit from highly proteinaceous unripe seeds to sugary fruit. They seem to avoid unripe and higher-fat fruit.
Variation in fruit consumption was positively associated with variation in fruit production. The gorillas ate fig fruits frequently; fig intake is positively correlated with that of other fruits, and figs were not fallback foods. They relied heavily on bamboo shoots on a seasonal basis; however, no bamboo shoots were available for several years after a major flowering event. Our results support the argument that variation in gorilla diets mostly reflects variation in vegetational composition of their habitats.
Despite the Bwindi gorillas having slightly lower availability of major food items, more fruit in their diets, and having longer daily travel distances than the Virunga gorillas, the energy intake rates for both populations were similar throughout the year. In Bwindi, when the gorillas were more frugivorous, their intake of protein declined and of carbohydrates was higher than that of Virunga gorillas.
Mountain gorillas prioritize consumption of non-protein energy sources (fruit) when available, yet select foods from their habitat that are high in protein.
When fruits are in season, western gorillas will spend more time foraging for fruits. Meanwhile, in the dry season, they will eat more leaves, stems, and other low-quality vegetable matter. Their preferred foods include fruits such as berries, bananas, and guavas. Over the course of a day, it’s not uncommon for a gorilla to consume 20 to 40 different types of foods.
We conclude that gorillas exploit the broad frugivore niche in West African lowland forests, and are part of the frugivore community there. Gorilla diet at the Lope Reserve overlaps greatly with that of sympatric, frugivorous, primates, and resembles more closely that of chimpanzees than it does gorilla diet studied elsewhere in Africa
The resultant life tables are truncated at age 55, the oldest age estimate reported for an individual from any of the five sites. Although some sites show temporal heterogeneity in mortality, we have chosen to include all time periods and all deaths reported at each site. The data include viral epidemics (polio and respiratory epidemics at Gombe, Ebola and Taï, influenza and an ‘‘Aids-like’’ epidemic at Mahale), deaths from poaching, warfare, infanticide, cannibalism and predation. Infant mortality is about 20% in the first year, dropping to a minimum of about 3·5% between ages 10–15. At age 15 the life expectancy is about another 15 years. By age 30 the annual mortality rate is about 8·5% and life expectancy an additional eight years.
Only 27% of all males born, but 41% of all females born are expected to survive to age 15
Muller and Wrangham argued that variation in pathogen exposure and in types and extent of anthropogenic disturbance largely explained inter-site variation in chimpanzee mortality and that the highest mortality rates (at Taï and Mahale) in the four-site sample reflected recent catastrophic human impacts. Ngogo data, which were not then available, are consistent with this argument. Exposure to pathogens carried by humans is relatively low at Ngogo and is probably lower than at Kanyawara, where the chimpanzees often raid crops (Wilson et al., 2014). Also, the Ngogo study area has suffered relatively little recent human disturbance. In fact, considerable forest regeneration in former anthropogenic grasslands has occurred in the Ngogo chimpanzee community territory since we began our study.
Life expectancy at birth (e0) for both sexes combined was 32.8 years. It was higher for females (35.8) than for males (29.6); this sex difference remained throughout life. Life expectancy at birth for Ngogo chimpanzees falls within the range of human hunter-gatherers and is similar to values for the Hadza, for whom life expectancy at birth is 35.5 for females and 30.8 for males. At 5 years of age, by which time Ngogo infants are typically weaned, future life expectancy was 36.5 years for the combined sample, 41.9 years for females, and 31 years for males.
These increases relative to life expectancy at birth are due to the concentration of early mortality in year 1. Values for females declined steadily thereafter, with no steep drop at any age.
For top-chain animals, captivity does have an obvious toll on their health despite their higher lifespan. This is due to medicine, otherwise they would die far younger from all kinds of ailments than their native counterparts. Just we live today longer than primitive hunter-gatherers, despite their higher constitutive health.
The most common causes of mortality were: cardiomyopathy (40% of all mortalities); stillbirth/abortion; acute myocardial necrosis; chimpanzee-induced trauma; amyloidosis; and pneumonia. Five morphologic diagnosis accounted for 61% of mortalities: cardiomyopathy; hemorrhage; acute myocardial necrosis; amyloidosis; and pneumonia. The most common etiologies were degenerative, undetermined, bacterial, traumatic, and neoplastic.
The cardiovascular system was most frequently involved, followed by the gastrointestinal, respiratory, and multi-systemic diseases.
The body of male WB, who died at the age of 26 in 2014, was found; he had been suffering from a respiratory infection that was the presumed cause of death. A few individuals who died around the same time also had respiratory infections, and some of those (e.g., male BE, 36 years old) looked weak and emaciated when last seen. Other chimpanzees were noticeably ill when last seen, and we assume that disease and/or the general effects of senescence are common sources of mortality. Male GZ apparently died from a fall because his body was found at the base of a tree e and D. Watts and J. Mitani saw an infant fall to its death as its mother fled from a male who was charging in the top of the canopy. Three females have been seen carrying dead infants presumed to have died of disease, but observers were unable to recover the bodies. Five infants (three of them first-born and two second-born to their respective mothers) have been victims of within-community infanticides, and one was a suspected victim of between-community infanticide.
We show that if one considers that clouds tend to form more frequently over forested areas, then planting trees over large areas is advantageous and should be done for climate purposes.
Princeton's Professor Amilcare Porporato
To reach this conclusion, researchers used satellite records of 2001-2010 cloud cover at latitudes of 30-45, comparing conditions in areas where forests had been replanted or planted for the first time. Not only do forests attract more cloud cover, they found, but clouds form above them earlier in the day, increasing the amount of sunlight they reflect.
Nevertheless, the authors acknowledge the competition that can take place with other land-uses, particularly agriculture.
Anthrax is rarely lethal for vultures or mammalian carnivores (Robertson 1976) although leopard (Panthera pardus), lion (P. leo), cheetah (Aciconyx jubatus), and genet (Genetta sp.) have been known to succumb to the disease under natural conditions (Pienaar 1960, 1961).
A special problem is that cattle may appear to recover completely from FMD but a number of them become carriers, and hence foci for new infection, for a long time (Bachrach 1978). Influenza virus seems to disappear from the human population after a while but is probably maintained in animal reservoirs (Kilbourne 1978); many migratory bird species contain this virus in their tissues (Lvov 1978). Polio, the third virus related to rinderpest, can survive for significant periods of time in the free state in water.
The means of transmission of these related virus “species” make it likely that, for example buffalo, can acquire rinderpest easily from the environment or that they can become infected through healthy carriers traveling from area to area.
Sinclair (1979) mentions that in the Serengeti, rinderpest tends to take its toll at the end of the dry season. Anthrax also tends to occur at the end of the dry season in the Kruger National Park (Pienaar 1961, 1967)?
The chance to get anthrax is dependent on ecological conditions [which] tend to occur at the end of the dry season.
Epidemics in Populations of Wild Ruminants: Anthrax and Impala, Rinderpest and Buffalo↩︎
Of all the regions of Kruger, the vegetation in the north is the least diversified and much of the region is blanketed in shrub mopane (Colophospermum mopane). The distribution of this tree in South Africa, Mozambique and Zimbabwe coincides with hot, semi-arid, low-lying valleys, and the mopane thrives under these conditions. Mopane leaves hang vertically and during the heat of the day very little shade is cast, which helps to minimize evaporation.
During the severe drought of the mid-1940s, the Letaba stopped flowing for two short spells at the end of winter. The Letaba and Olifants rivers are home to 60 per cent of the Park’s hippo, and in the past large numbers of hippo died in times of drought.
Other sites don’t show nearly that amount of violence, and much altruistic behaviors completely contradicting the notion of a mean ape always on the lookout for reproductive opportunities. Many of the data on wild-chimpanzee interactions come from East Africa, but troops living in the Taï forest, in Ivory Coast, clash less frequently; their conflicts are not so brutal and the females are subject to less of a power differential. Male chimps at West African sites have also been seen adopting the dependent young of a deceased or missing parent, a shouldering of fatherly duty at odds with newborn killings witnessed elsewhere.
In contrast, most bacteria in the phylum Firmicutes have a single membrane covered by a thick layer of peptidoglycan. The phylum Actinobacteria is quite distinct from the classical types, their thin peptidoglycan layer is directly covered by a thick polysaccharide layer. Many bacteria also contain a protein S-layer. ↩︎
One can imagine a molecular imaging system derived from MS2-GFP and trimolecular fluorescence complementation (TriFC), where the quench (signal quenching element of the fluorescent construct) would only detach/activate, when RAGE and an AGE would be present, or reporters covalently fused with AGEs in solution. ↩︎